

Epistasis studies reveal redundancy among calcium-dependent protein kinases in motility and invasion of malaria parasites
- PMID: 30315162
- PMCID: PMC6185908
- DOI: 10.1038/s41467-018-06733-w
Epistasis studies reveal redundancy among calcium-dependent protein kinases in motility and invasion of malaria parasites
Abstract
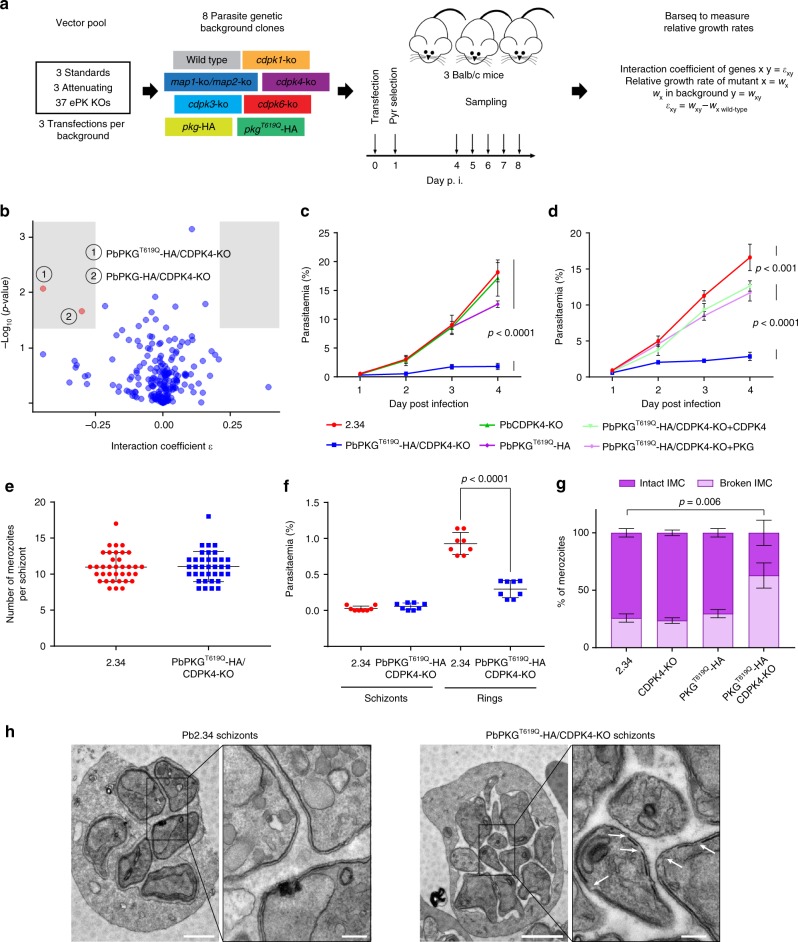
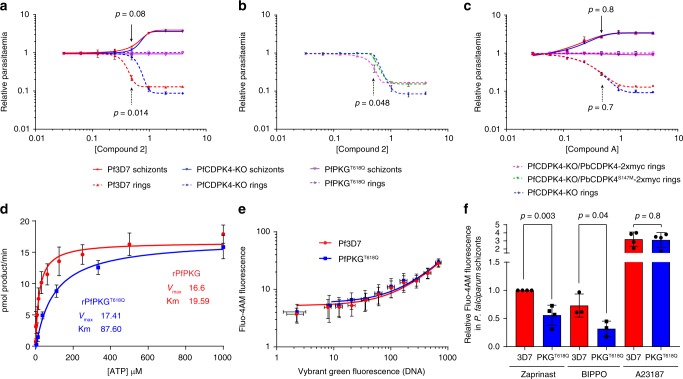
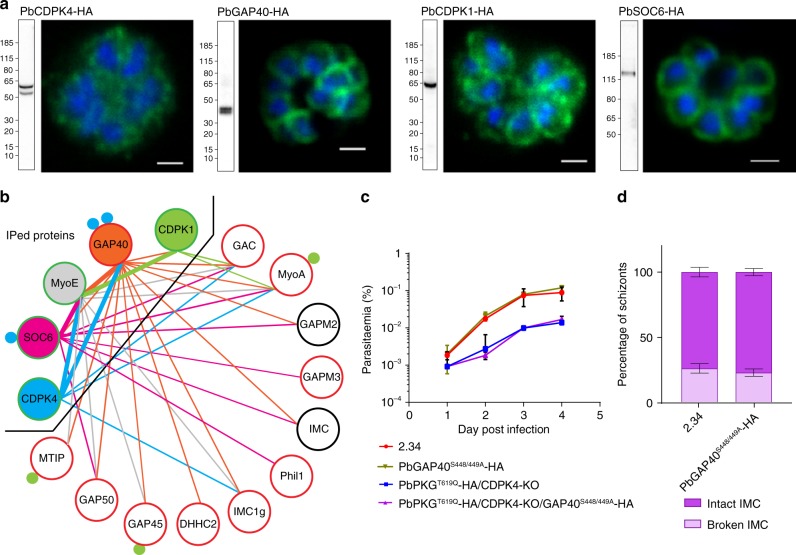
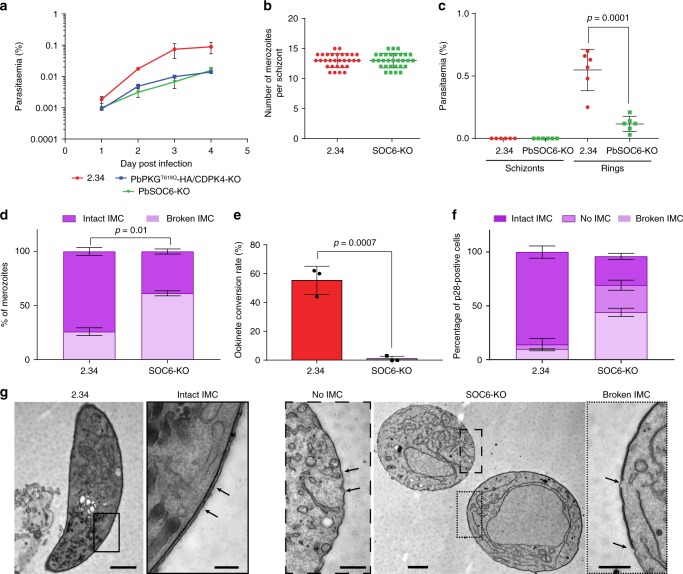
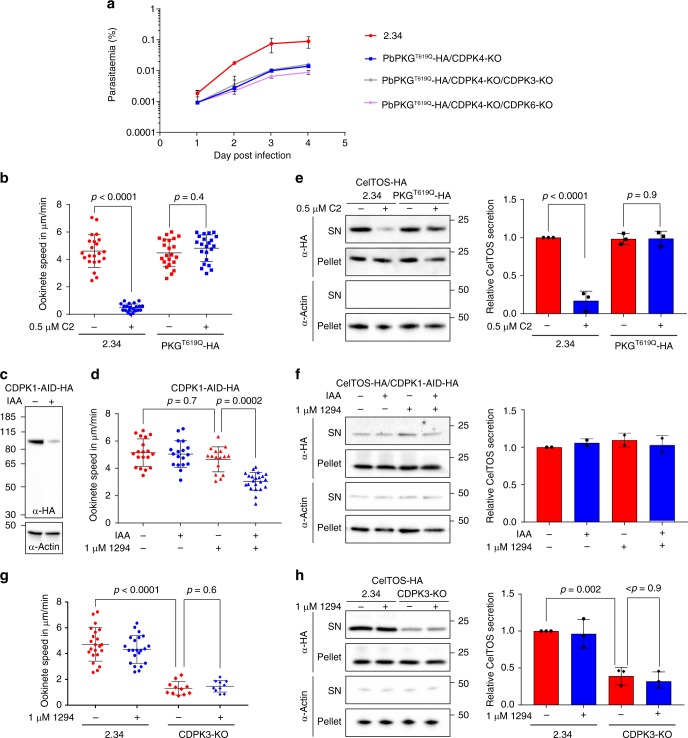
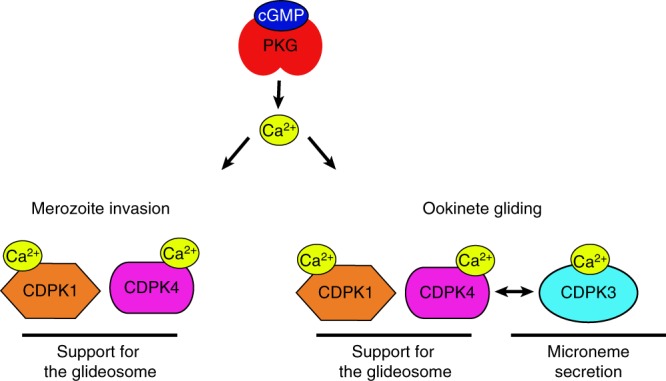
In malaria parasites, evolution of parasitism has been linked to functional optimisation. Despite this optimisation, most members of a calcium-dependent protein kinase (CDPK) family show genetic redundancy during erythrocytic proliferation. To identify relationships between phospho-signalling pathways, we here screen 294 genetic interactions among protein kinases in Plasmodium berghei. This reveals a synthetic negative interaction between a hypomorphic allele of the protein kinase G (PKG) and CDPK4 to control erythrocyte invasion which is conserved in P. falciparum. CDPK4 becomes critical when PKG-dependent calcium signals are attenuated to phosphorylate proteins important for the stability of the inner membrane complex, which serves as an anchor for the acto-myosin motor required for motility and invasion. Finally, we show that multiple kinases functionally complement CDPK4 during erythrocytic proliferation and transmission to the mosquito. This study reveals how CDPKs are wired within a stage-transcending signalling network to control motility and host cell invasion in malaria parasites.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Brochet M., Tobin A. B., Billker O. & Doerig C. in Kinomics: Approaches and Applications (eds Kraatz, H.-B. & Martic, S.) 115–136 (John Wiley & Sons, Weinheim, 2015).
Publication types
MeSH terms
Substances
Grants and funding
- 695596/EC | European Research Council (ERC)/International
- G1000779/MRC_/Medical Research Council/United Kingdom
- 106240/Z/14/Z/WT_/Wellcome Trust/United Kingdom
- BSSGI0_155852/Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (Swiss National Science Foundation)/International
- 098051/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
