

The nonsense-mediated mRNA decay (NMD) pathway differentially regulates COX17, COX19 and COX23 mRNAs
- PMID: 30317392
- PMCID: PMC6420912
- DOI: 10.1007/s00294-018-0892-y
The nonsense-mediated mRNA decay (NMD) pathway differentially regulates COX17, COX19 and COX23 mRNAs
Abstract
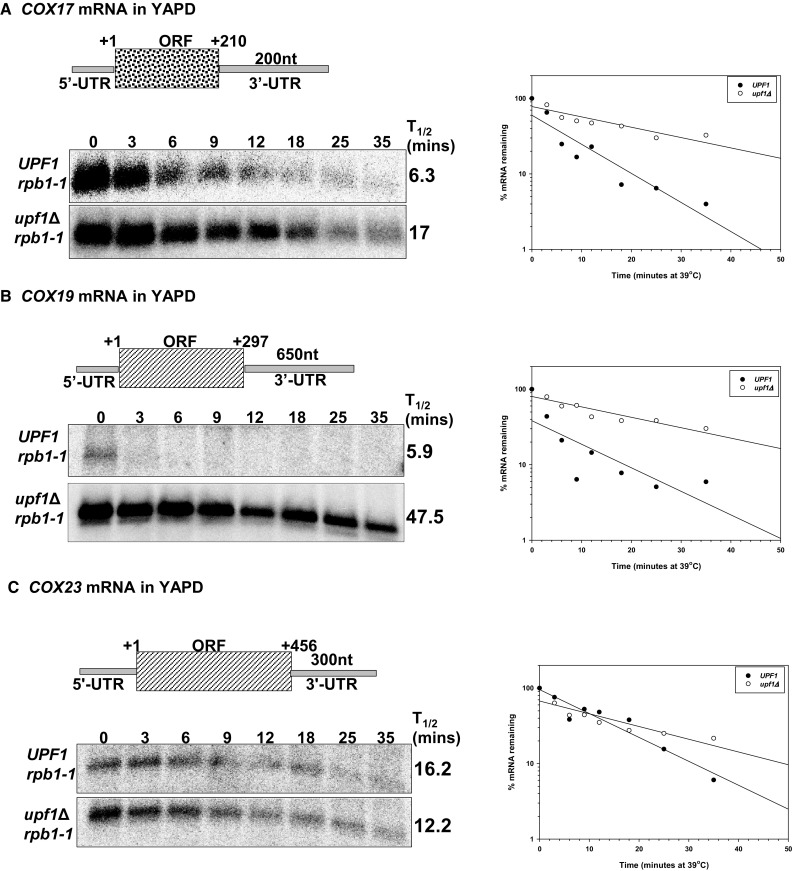
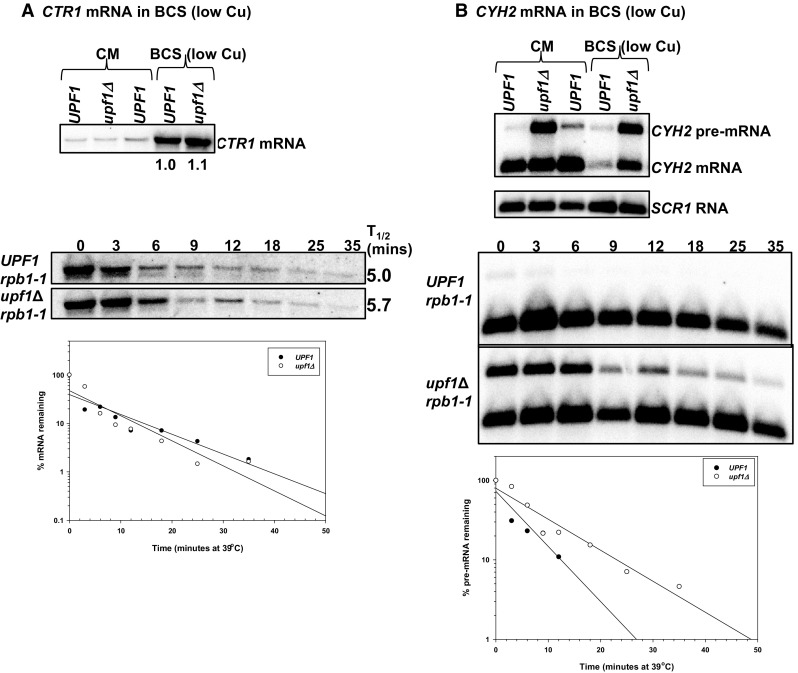
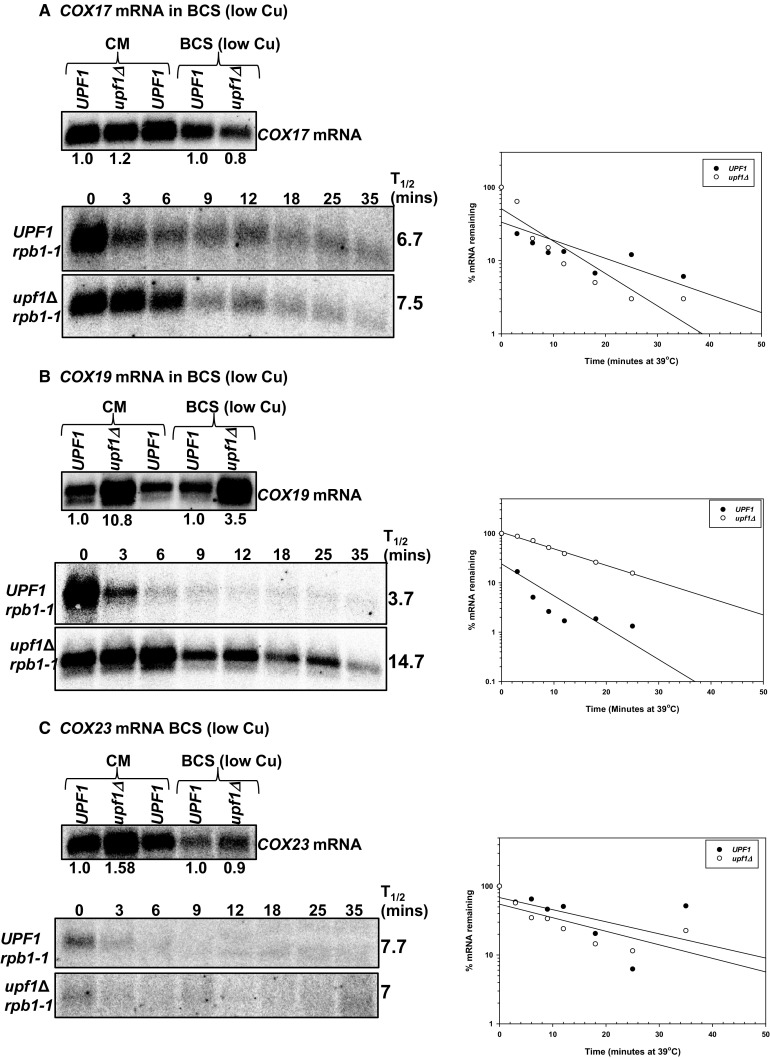
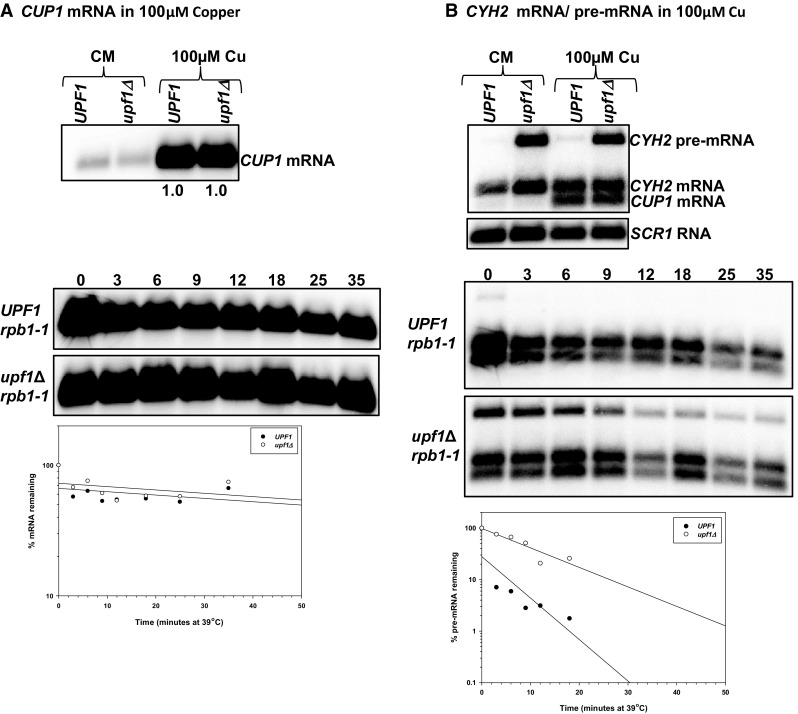
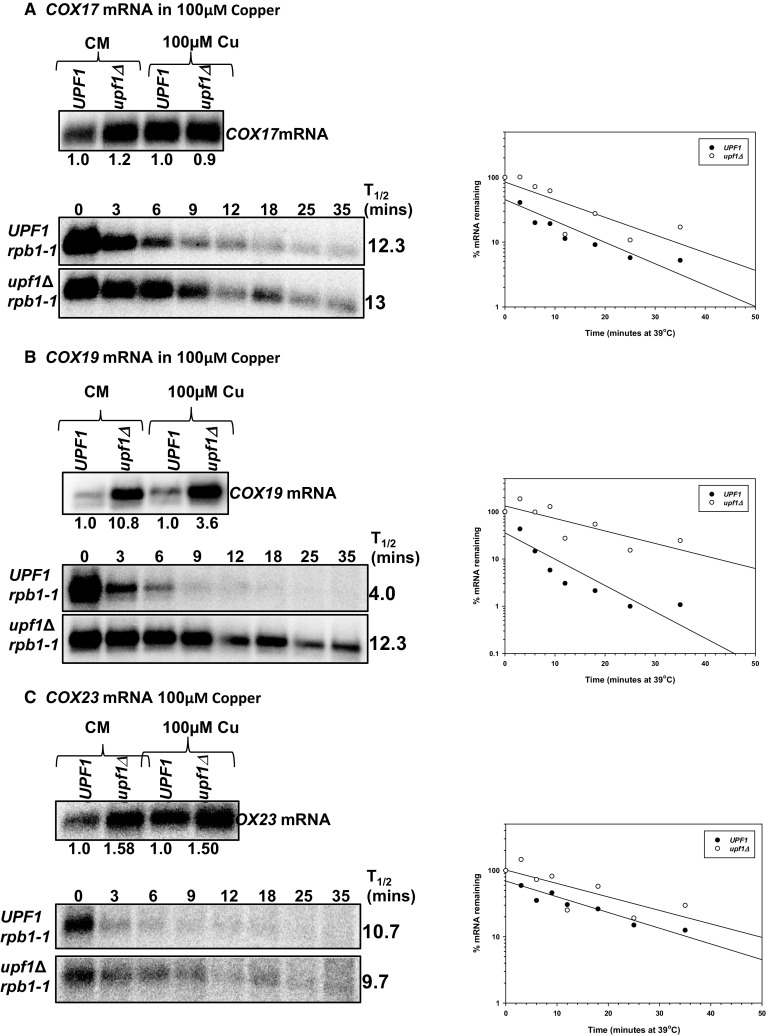
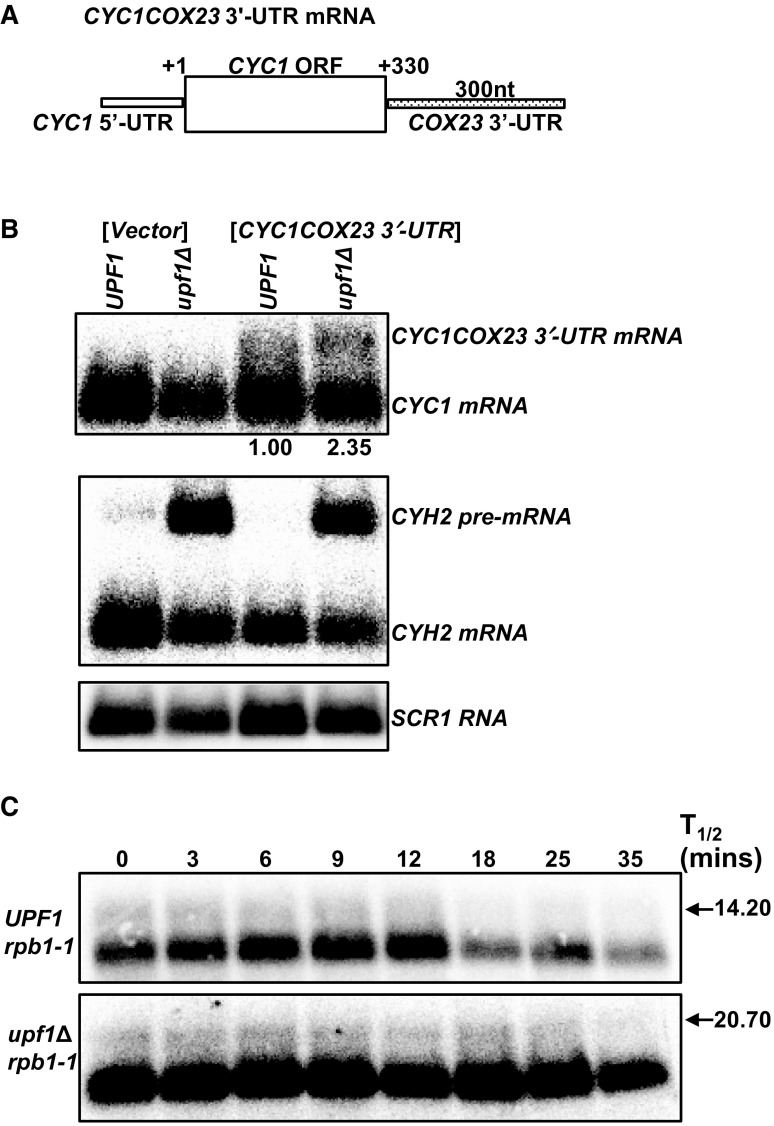
The differential regulation of COX17, COX19 and COX23 mRNAs by the nonsense-mediated mRNA decay (NMD) pathway was investigated. The NMD pathway regulates mRNAs that aberrantly terminate translation. This includes mRNAs harboring premature translation termination codons and natural mRNAs. Most natural mRNAs regulated by NMD encode fully functional proteins involved in various cellular processes. However, the cause and targeting of most of these mRNAs by the pathway is not understood. Analysis of a set of mRNAs involved in copper homeostasis showed that a subset of these mRNAs function in mitochondrial copper homeostasis. Here, we examined the regulation of COX17, COX19 and COX23 mRNAs by NMD. These mRNAs encode homologous mitochondrial proteins involved in metallation of cytochrome c oxidase. We found that COX17, COX19 and COX23 mRNAs are differentially regulated by NMD depending on environmental copper levels. A long 3'-UTR contributes to the direct regulation of COX19 mRNA by the pathway. Alternatively, COX23 mRNA contains a long 3'-UTR, but is indirectly regulated by the pathway under two conditions tested here. Analysis of the functionality of the NMD targeting features in COX23 mRNA showed that the COX23 3'-UTR is sufficient to trigger NMD. The regulation of mRNAs involved in mitochondrial copper metabolism by NMD is physiologically significant because excess copper enhances growth of NMD mutants on a non-fermentable carbon source. These findings suggest that regulation of mRNAs encoding homologous proteins by NMD can be differential depending on environmental copper levels. Furthermore, these findings suggest copper ion homeostatic mechanisms in the mitochondria occur at the mRNA level via the NMD pathway.
Keywords: Mitochondrial copper homeostasis; Nonsense-mediated mRNA decay; Saccharomyces cerevisiae; mRNA; mRNA decay.
Conflict of interest statement
The research presented in this manuscript has not been submitted previously or concurrently to another journal, does not contain any fabricated or manipulated data and has not been split into several parts. The contributions of each of the author listed are detailed above in the authors’ contributions.
Figures


References
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current protocols in molecular biology. New York: Wiley; 1998.
-
- De Paepe B, Lefever S, Mestdagh P. How long noncoding RNAs enforce their will on mitochondrial activity: regulation of mitochondrial respiration, reactive oxygen species production, apoptosis, and metabolic reprogramming in cancer. Curr Genet. 2018;64:163–172. doi: 10.1007/s00294-017-0744-1. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
