

The Adrenal Clock Prevents Aberrant Light-Induced Alterations in Circadian Glucocorticoid Rhythms
- PMID: 30321360
- PMCID: PMC6240903
- DOI: 10.1210/en.2018-00769
The Adrenal Clock Prevents Aberrant Light-Induced Alterations in Circadian Glucocorticoid Rhythms
Abstract
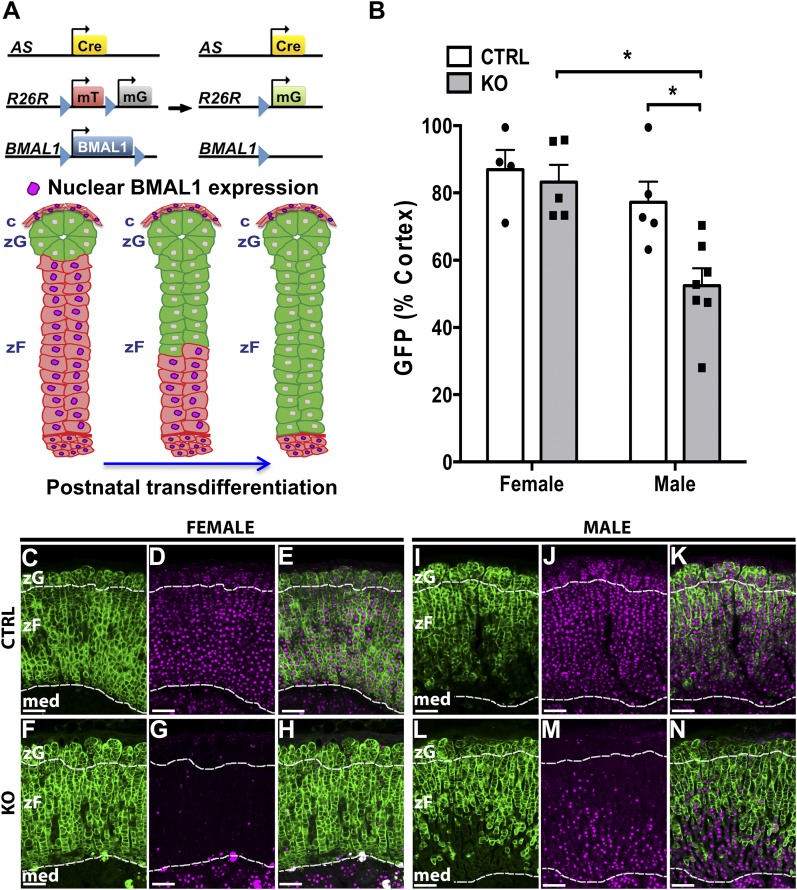
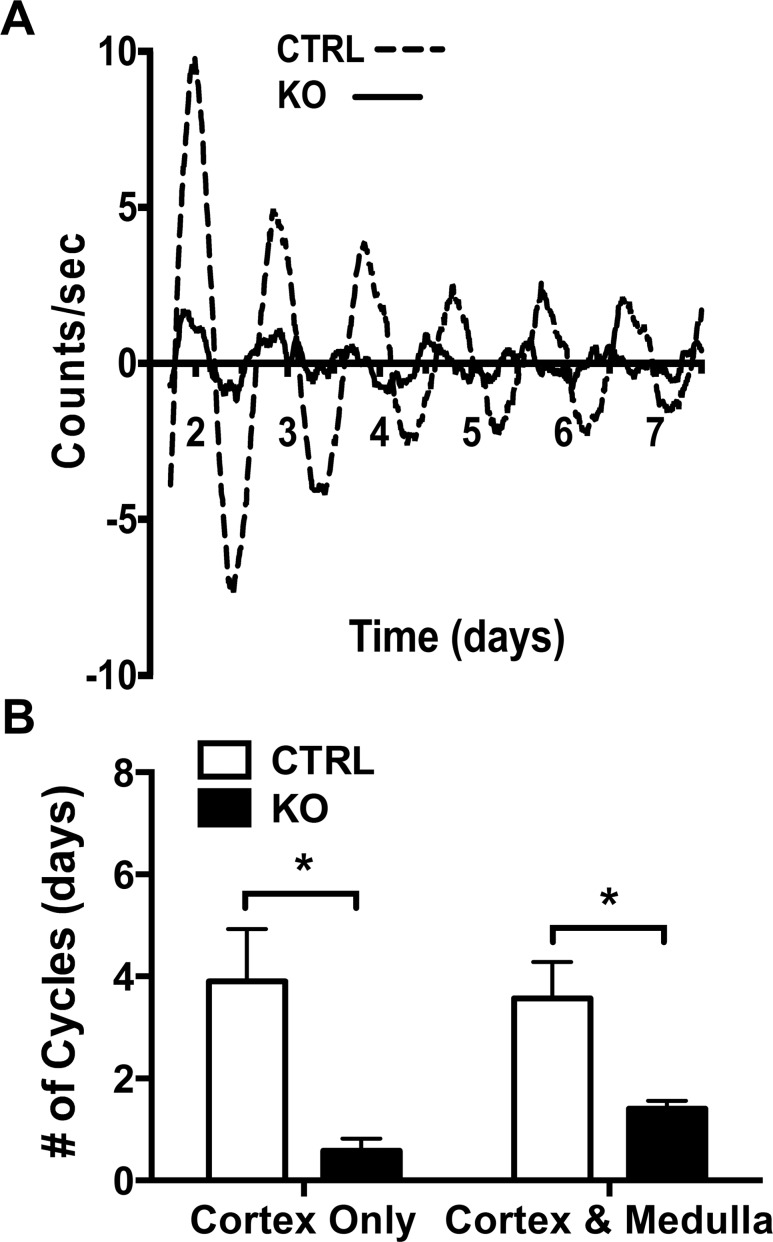
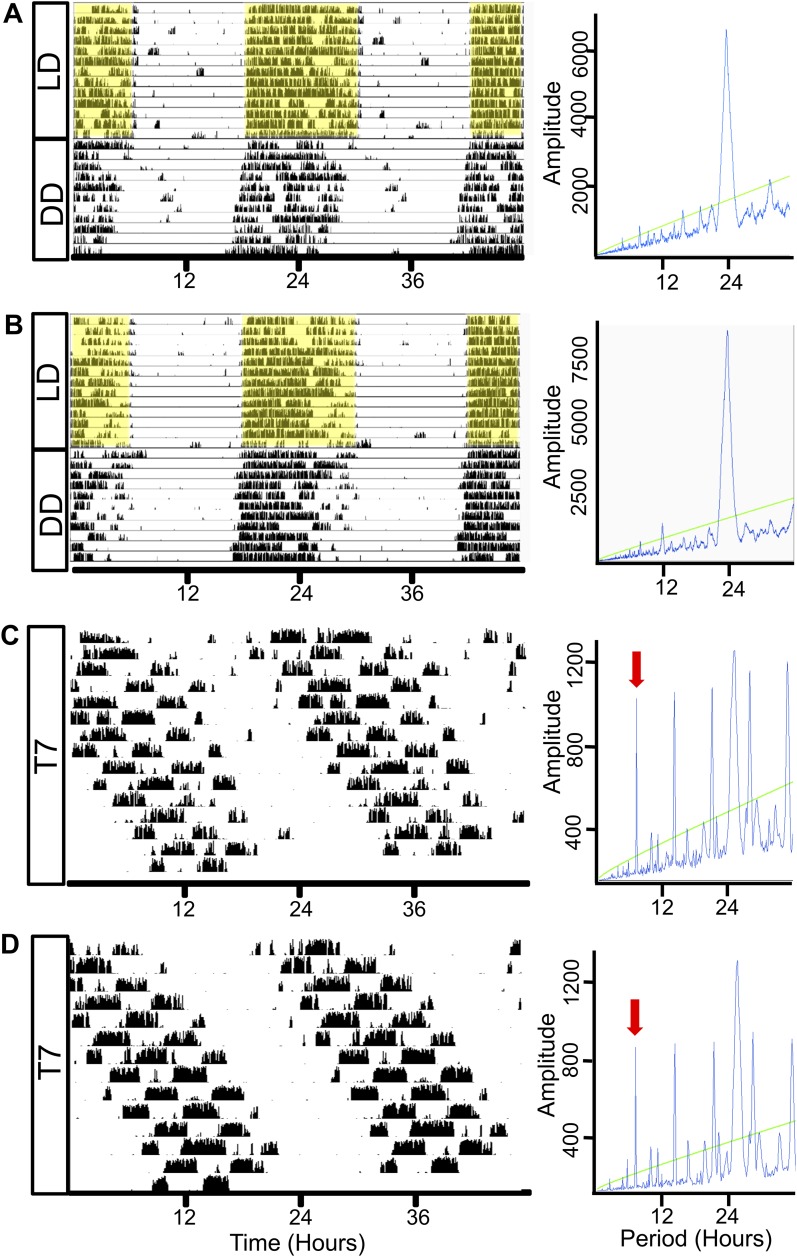
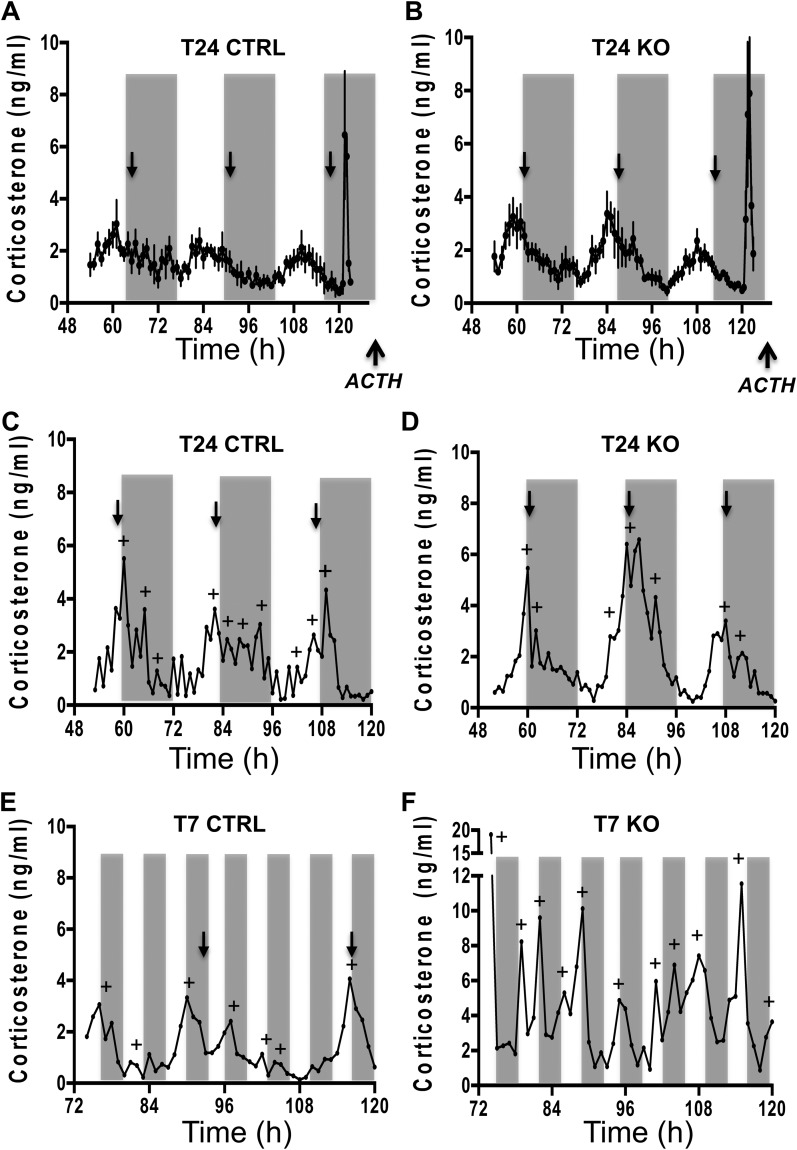
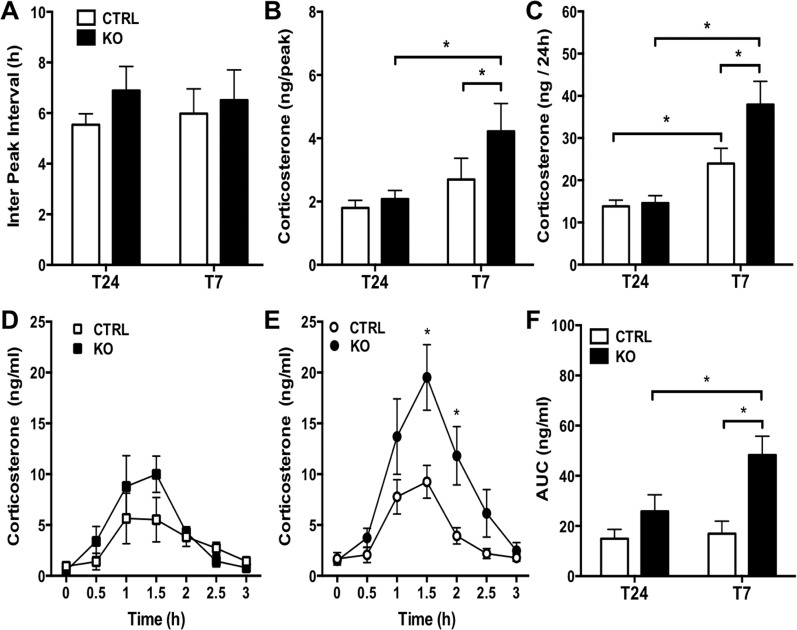
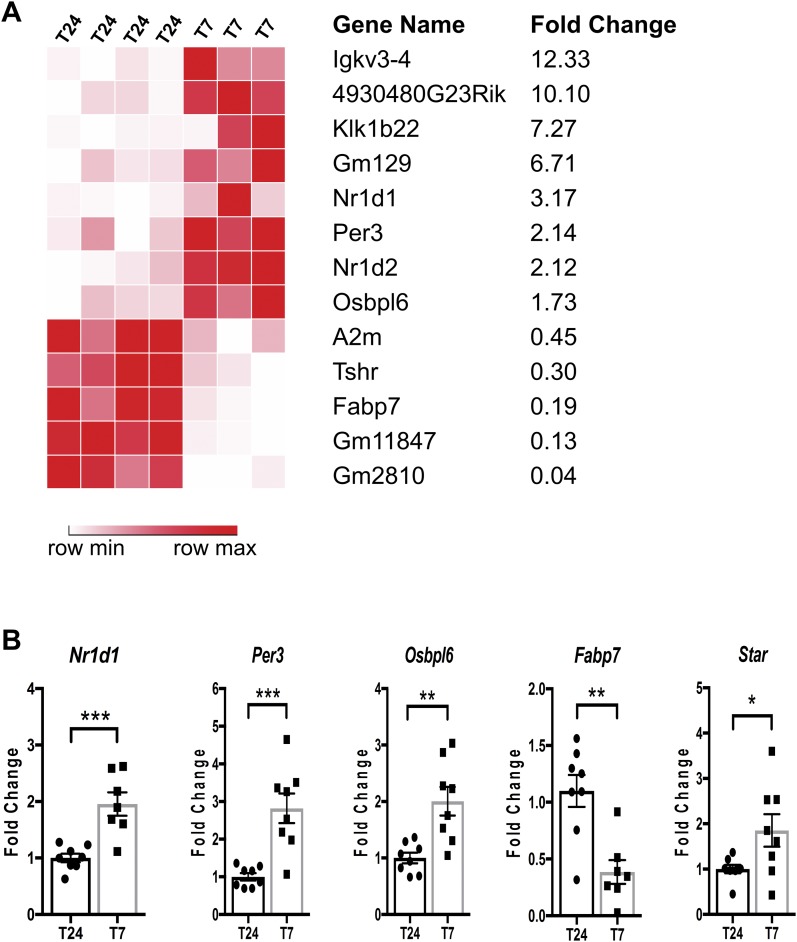
The glucocorticoid (GC) rhythm is entrained to light-dark (LD) cycles via a molecular clock in the suprachiasmatic nucleus (SCN) and is maintained by an adrenal clock synchronized by SCN-dependent signals. Targeted deletion of the core clock gene Bmal1 can disrupt adrenal clock function. The requirement of the adrenal clock to stabilize the circadian GC rhythm during exposure to aberrant LD cycles was determined using novel aldosterone synthase (AS)Cre/+::Bmal1Fl/Fl mice in which Bmal1 deletion occurred during postnatal adrenal transdifferentiation. To examine whether adrenal Bmal1 deletion results in loss of the adrenal clock, mice were crossed with mPER2::Luciferase (mPER2Luc/+) mice. Adrenals from ASCre/+::Bmal1+/+::PER2Luc/+ [control (CTRL)] mice show mPER2Luc rhythms ex vivo, whereas slices from ASCre/+::Bmal1Fl/Fl::PER2Luc/+ [knockout (KO)] mice show dampened rhythms. To monitor corticosterone rhythmicity, mice were implanted with subcutaneous microdialysis probes and sampled at 60-minute intervals for up to 3 days under 12:12-hour [τ (T) 24] LD or 3.5:3.5-hour (T7) LD cycles. Corticosterone rhythms were entrained to T24 LD in CTRL and KO mice. Under T7 LD, circadian corticosterone rhythms persisted in most CTRL mice but not KO mice. Hyperadrenocorticism also was observed in KO mice under T7 LD, reflected by increased corticosterone peak amplitude, total daily corticosterone, and responses to ACTH. Analysis of dysregulated adrenal genes in KO mice exposed to aberrant light identified candidates involved in cholesterol metabolism and trafficking, including steroidogenic acute regulatory protein, which could increase steroidogenesis. Our results show that the adrenal clock functions to buffer steroidogenic responses to aberrant light and stabilize circadian GC rhythmicity.
Figures
References
-
- Zelinski EL, Deibel SH, McDonald RJ. The trouble with circadian clock dysfunction: multiple deleterious effects on the brain and body. Neurosci Biobehav Rev. 2014;40:80–101. - PubMed
-
- Reppert SM, Weaver DR. Coordination of circadian timing in mammals. Nature. 2002;418(6901):935–941. - PubMed
-
- Dibner C, Schibler U, Albrecht U. The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annu Rev Physiol. 2010;72(1):517–549. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
