

Human GTPBP10 is required for mitoribosome maturation
- PMID: 30321378
- PMCID: PMC6265488
- DOI: 10.1093/nar/gky938
Human GTPBP10 is required for mitoribosome maturation
Abstract
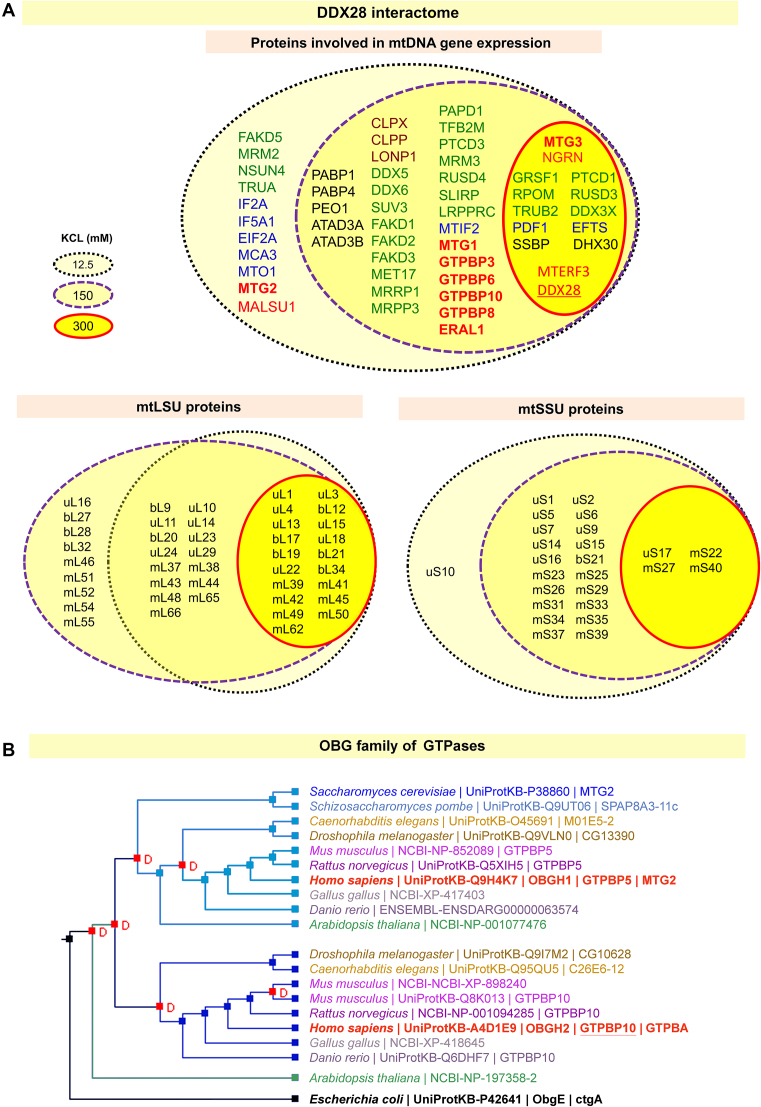
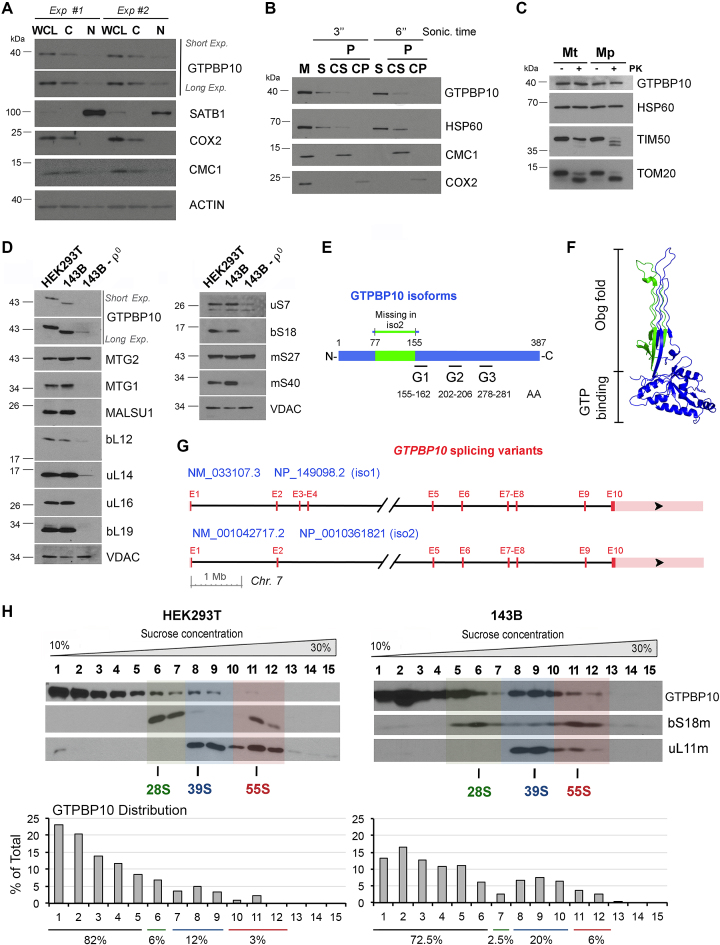
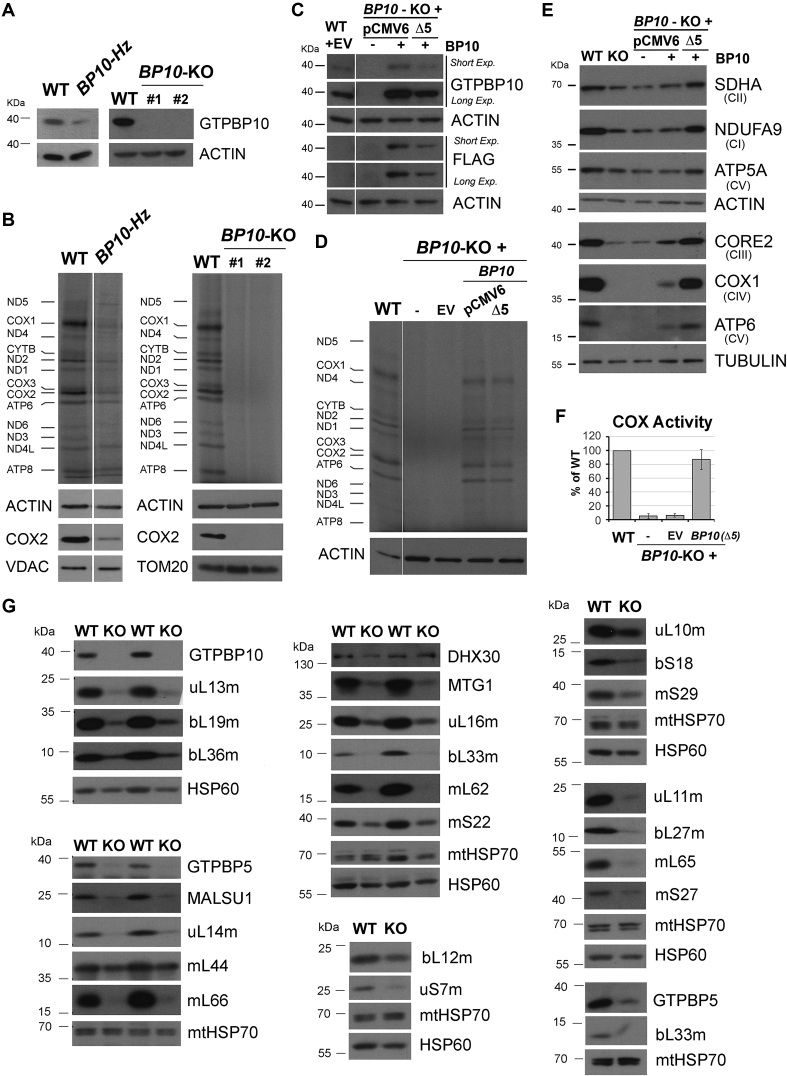
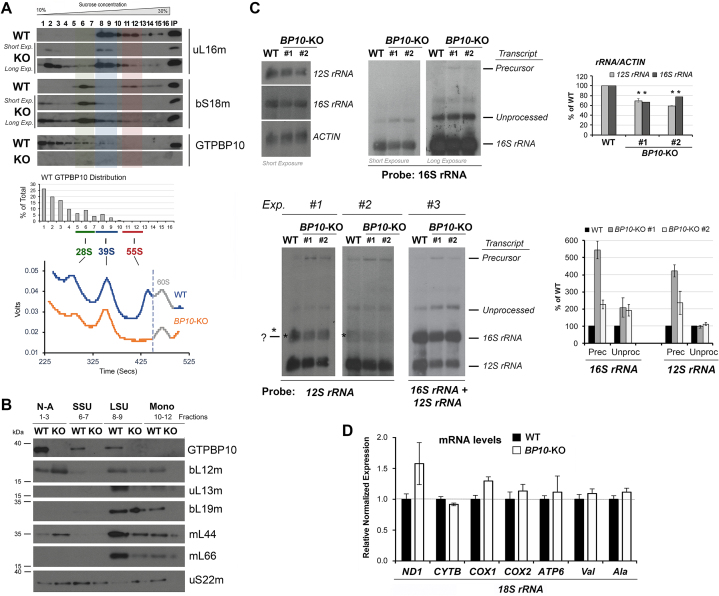
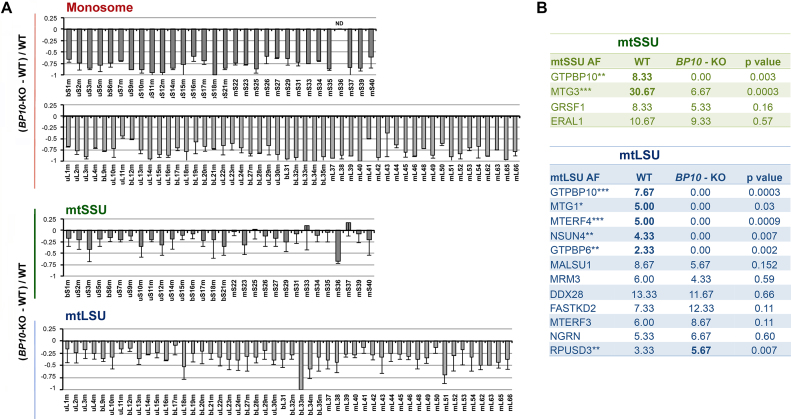
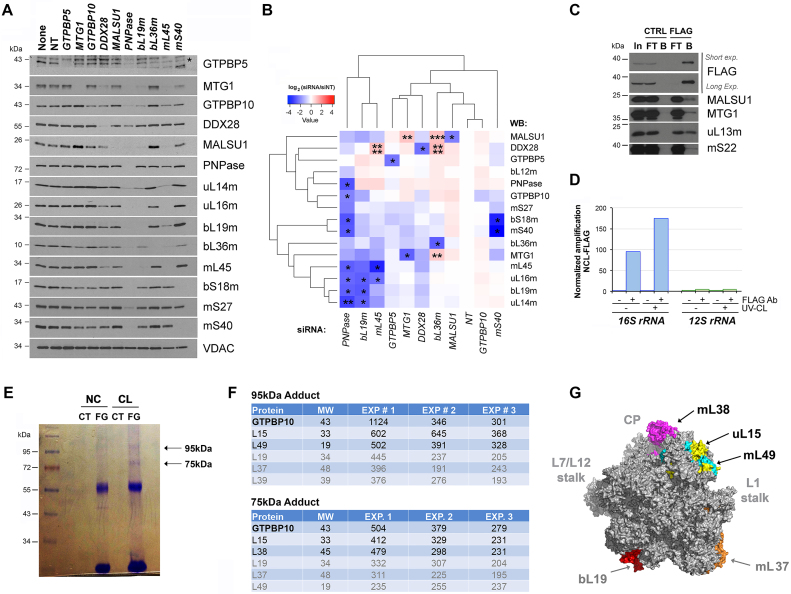
Most steps on the biogenesis of the mitochondrial ribosome (mitoribosome) occur near the mitochondrial DNA nucleoid, in RNA granules, which contain dedicated RNA metabolism and mitoribosome assembly factors. Here, analysis of the RNA granule proteome identified the presence of a set of small GTPases that belong to conserved families of ribosome assembly factors. We show that GTPBP10, a member of the conserved Obg family of P-loop small G proteins, is a mitochondrial protein and have used gene-editing technologies to create a HEK293T cell line KO for GTPBP10. The absence of GTPBP10 leads to attenuated mtLSU and mtSSU levels and the virtual absence of the 55S monosome, which entirely prevents mitochondrial protein synthesis. We show that a fraction of GTPBP10 cosediments with the large mitoribosome subunit and the monosome. GTPBP10 physically interacts with the 16S rRNA, but not with the 12S rRNA, and crosslinks with several mtLSU proteins. Additionally, GTPBP10 is indirectly required for efficient processing of the 12S-16S rRNA precursor transcript, which could explain the mtSSU accumulation defect. We propose that GTPBP10 primarily ensures proper mtLSU maturation and ultimately serves to coordinate mtSSU and mtLSU accumulation then providing a quality control check-point function during mtLSU assembly that minimizes premature subunit joining.
Figures
References
-
- Greber B.J., Bieri P., Leibundgut M., Leitner A., Aebersold R., Boehringer D., Ban N.. Ribosome. The complete structure of the 55S mammalian mitochondrial ribosome. Science. 2015; 348:303–308. - PubMed
-
- Greber B.J., Boehringer D., Leibundgut M., Bieri P., Leitner A., Schmitz N., Aebersold R., Ban N.. The complete structure of the large subunit of the mammalian mitochondrial ribosome. Nature. 2014; 515:283–286. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
