

Monitoring Protein Dynamics in Protein O-Mannosyltransferase Mutants In Vivo by Tandem Fluorescent Protein Timers
- PMID: 30322079
- PMCID: PMC6222916
- DOI: 10.3390/molecules23102622
Monitoring Protein Dynamics in Protein O-Mannosyltransferase Mutants In Vivo by Tandem Fluorescent Protein Timers
Abstract
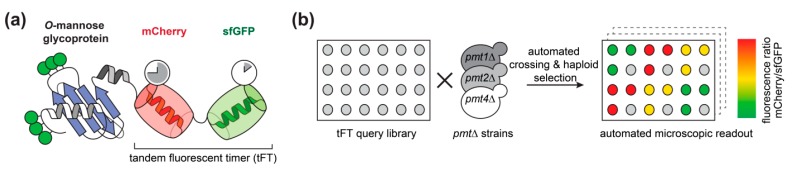
For proteins entering the secretory pathway, a major factor contributing to maturation and homeostasis is glycosylation. One relevant type of protein glycosylation is O-mannosylation, which is essential and evolutionarily-conserved in fungi, animals, and humans. Our recent proteome-wide study in the eukaryotic model organism Saccharomyces cerevisiae revealed that more than 26% of all proteins entering the secretory pathway receive O-mannosyl glycans. In a first attempt to understand the impact of O-mannosylation on these proteins, we took advantage of a tandem fluorescent timer (tFT) reporter to monitor different aspects of protein dynamics. We analyzed tFT-reporter fusions of 137 unique O-mannosylated proteins, mainly of the secretory pathway and the plasma membrane, in mutants lacking the major protein O-mannosyltransferases Pmt1, Pmt2, or Pmt4. In these three pmtΔ mutants, a total of 39 individual proteins were clearly affected, and Pmt-specific substrate proteins could be identified. We observed that O-mannosylation may cause both enhanced and diminished protein abundance and/or stability when compromised, and verified our findings on the examples of Axl2-tFT and Kre6-tFT fusion proteins. The identified target proteins are a valuable resource towards unraveling the multiple functions of O-mannosylation at the molecular level.
Keywords: O-mannosyl glycans; PMT1; PMT2; PMT4; Saccharomyces cerevisiae; fluorescent protein timers; glycosylation; mannosyltransferase; protein turnover; secretory pathway; yeast.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Similar articles
-
Translational Regulation of Pmt1 and Pmt2 by Bfr1 Affects Unfolded Protein O-Mannosylation.Int J Mol Sci. 2019 Dec 10;20(24):6220. doi: 10.3390/ijms20246220. Int J Mol Sci. 2019. PMID: 31835530 Free PMC article.
-
O-mannosylation precedes and potentially controls the N-glycosylation of a yeast cell wall glycoprotein.EMBO Rep. 2003 Jun;4(6):628-32. doi: 10.1038/sj.embor.embor864. EMBO Rep. 2003. PMID: 12776183 Free PMC article.
-
Protein-O-glycosylation in yeast: protein-specific mannosyltransferases.Glycobiology. 1997 Jun;7(4):481-6. doi: 10.1093/glycob/7.4.481. Glycobiology. 1997. PMID: 9184828
-
Protein O-mannosylation: what we have learned from baker's yeast.Biochim Biophys Acta. 2013 Nov;1833(11):2438-46. doi: 10.1016/j.bbamcr.2013.02.008. Epub 2013 Feb 20. Biochim Biophys Acta. 2013. PMID: 23434682 Review.
-
Protein O-mannosylation.Biochim Biophys Acta. 1999 Jan 6;1426(2):297-307. doi: 10.1016/s0304-4165(98)00131-7. Biochim Biophys Acta. 1999. PMID: 9878797 Review.
Cited by
-
On the Potential Function of Type II Arabinogalactan O-Glycosylation in Regulating the Fate of Plant Secretory Proteins.Front Plant Sci. 2020 Sep 10;11:563735. doi: 10.3389/fpls.2020.563735. eCollection 2020. Front Plant Sci. 2020. PMID: 33013983 Free PMC article. Review.
-
Blue-to-Red TagFT, mTagFT, mTsFT, and Green-to-FarRed mNeptusFT2 Proteins, Genetically Encoded True and Tandem Fluorescent Timers.Int J Mol Sci. 2023 Feb 7;24(4):3279. doi: 10.3390/ijms24043279. Int J Mol Sci. 2023. PMID: 36834686 Free PMC article.
-
Translational Regulation of Pmt1 and Pmt2 by Bfr1 Affects Unfolded Protein O-Mannosylation.Int J Mol Sci. 2019 Dec 10;20(24):6220. doi: 10.3390/ijms20246220. Int J Mol Sci. 2019. PMID: 31835530 Free PMC article.
-
Functional Roles of O-Glycosylation.Molecules. 2018 Nov 23;23(12):3063. doi: 10.3390/molecules23123063. Molecules. 2018. PMID: 30477085 Free PMC article.
References
-
- Sentandreu R., Northcote D.H. The characterization of oligosaccharide attachment to threonine and serine in mannan glycopeptides obtained from the cell wall of yeast. Carbohydr. Res. 1969;10:584–585. doi: 10.1016/S0008-6215(00)80126-3. - DOI
-
- Larsen I.S.B., Narimatsu Y., Joshi H.J., Siukstaite L., Harrison O.J., Brasch J., Goodman K.M., Hansen L., Shapiro L., Honig B., et al. Discovery of an O-mannosylation pathway selectively serving cadherins and protocadherins. Proc. Natl. Acad. Sci. USA. 2017;114:11163–11168. doi: 10.1073/pnas.1708319114. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
