

Microtubules Provide a Viscoelastic Resistance to Myocyte Motion
- PMID: 30322798
- PMCID: PMC6224693
- DOI: 10.1016/j.bpj.2018.09.019
Microtubules Provide a Viscoelastic Resistance to Myocyte Motion
Abstract
Background: Microtubules (MTs) buckle and bear load during myocyte contraction, a behavior enhanced by post-translational detyrosination. This buckling suggests a spring-like resistance against myocyte shortening, which could store energy and aid myocyte relaxation. Despite this visual suggestion of elastic behavior, the precise mechanical contribution of the cardiac MT network remains to be defined.
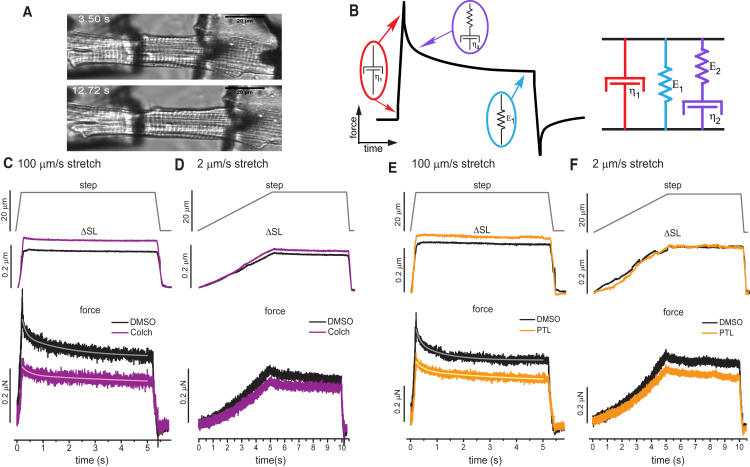
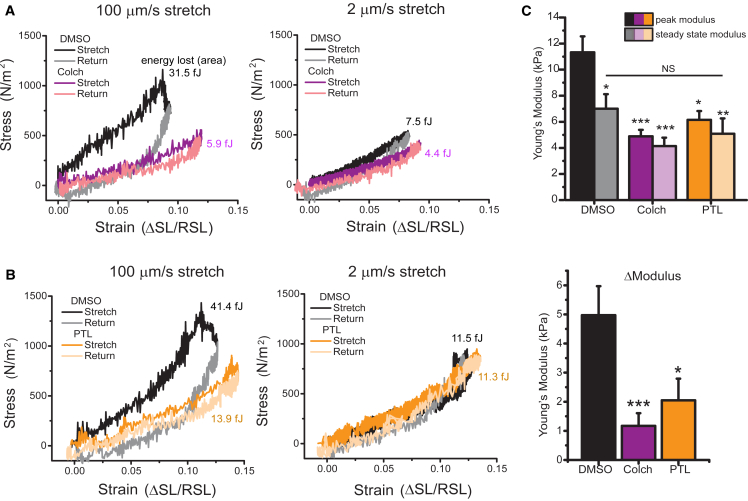
Methods: Here we experimentally and computationally probe the mechanical contribution of stable MTs and their influence on myocyte function. We use multiple approaches to interrogate viscoelasticity and cell shortening in primary murine myocytes in which either MTs are depolymerized or detyrosination is suppressed and use the results to inform a mathematical model of myocyte viscoelasticity.
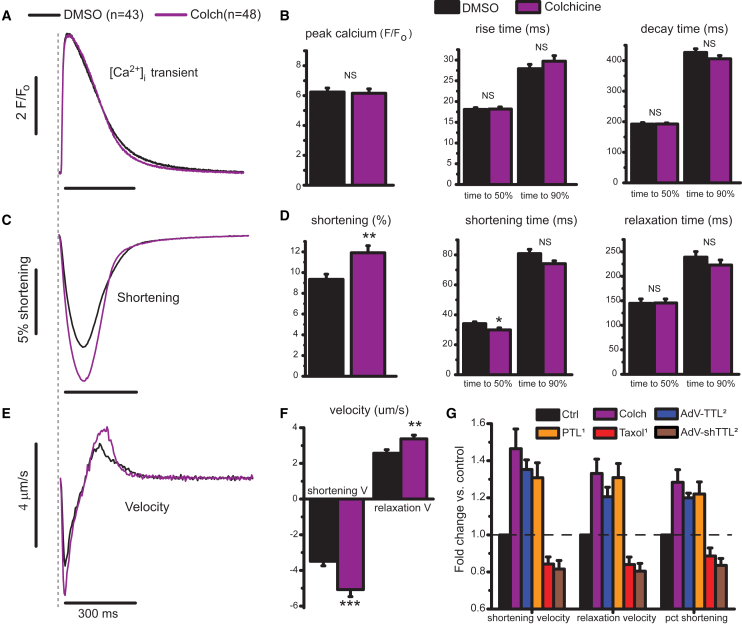
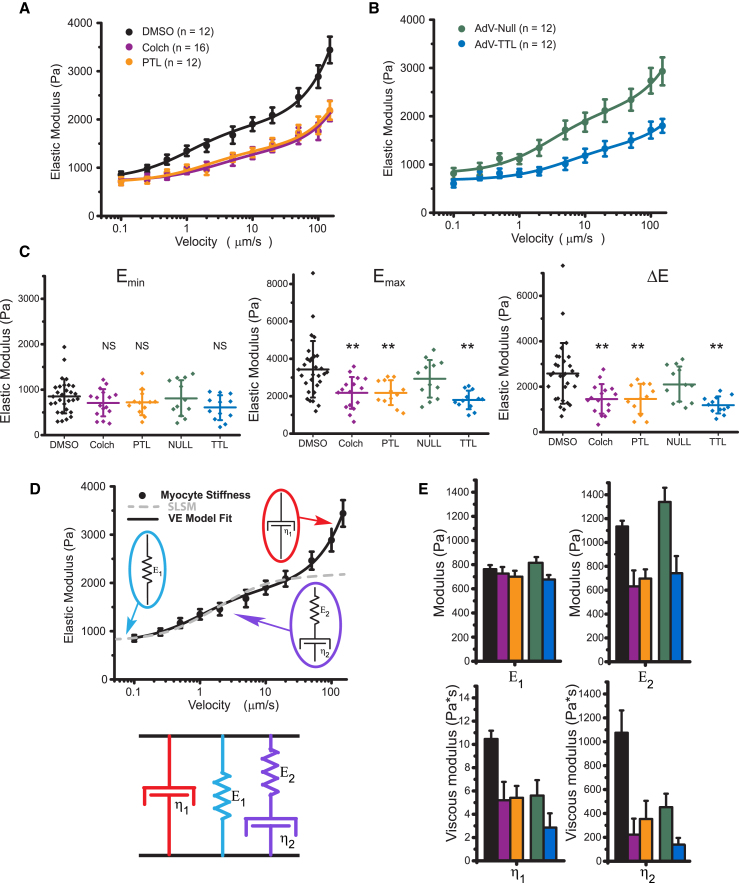
Results: MT ablation by colchicine concurrently enhances both the degree of shortening and speed of relaxation, a finding inconsistent with simple spring-like MT behavior and suggestive of a viscoelastic mechanism. Axial stretch and transverse indentation confirm that MTs increase myocyte viscoelasticity. Specifically, increasing the rate of strain amplifies the MT contribution to myocyte stiffness. Suppressing MT detyrosination with parthenolide or via overexpression of tubulin tyrosine ligase has mechanical consequences that closely resemble colchicine, suggesting that the mechanical impact of MTs relies on a detyrosination-dependent linkage with the myocyte cytoskeleton. Mathematical modeling affirms that alterations in cell shortening conferred by either MT destabilization or tyrosination can be attributed to internal changes in myocyte viscoelasticity.
Conclusions: The results suggest that the cardiac MT network regulates contractile amplitudes and kinetics by acting as a cytoskeletal shock-absorber, whereby MTs provide breakable cross-links between the sarcomeric and nonsarcomeric cytoskeleton that resist rapid length changes during both shortening and stretch.
Copyright © 2018 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Janke C., Bulinski J.C. Post-translational regulation of the microtubule cytoskeleton: mechanisms and functions. Nat. Rev. Mol. Cell Biol. 2011;12:773–786. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
