

On the deformability of an empirical fitness landscape by microbial evolution
- PMID: 30322921
- PMCID: PMC6217403
- DOI: 10.1073/pnas.1808485115
On the deformability of an empirical fitness landscape by microbial evolution
Abstract
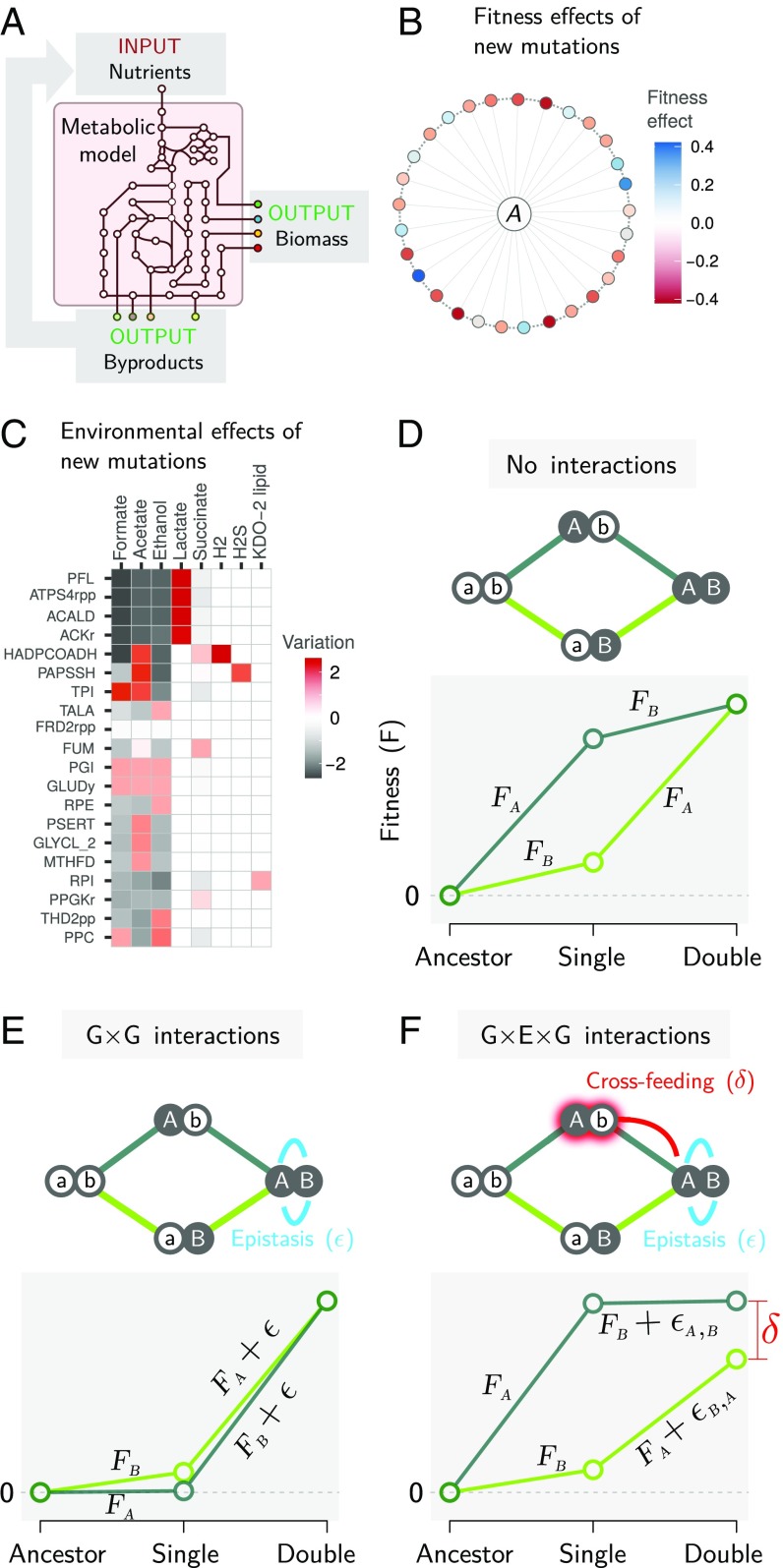
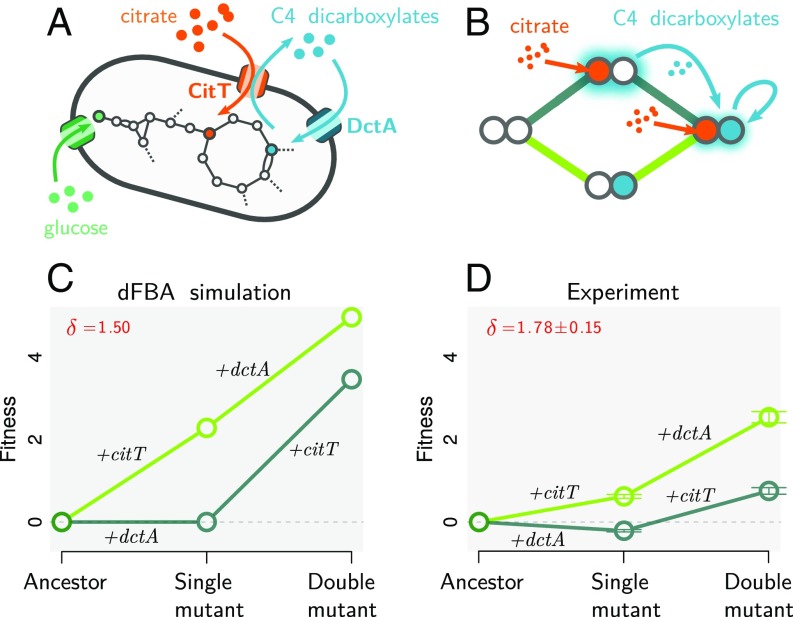
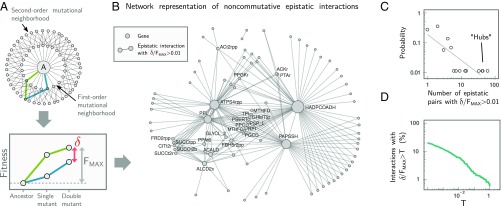
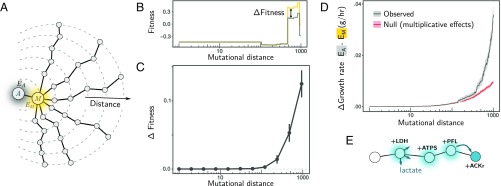
A fitness landscape is a map between the genotype and its reproductive success in a given environment. The topography of fitness landscapes largely governs adaptive dynamics, constraining evolutionary trajectories and the predictability of evolution. Theory suggests that this topography can be deformed by mutations that produce substantial changes to the environment. Despite its importance, the deformability of fitness landscapes has not been systematically studied beyond abstract models, and little is known about its reach and consequences in empirical systems. Here we have systematically characterized the deformability of the genome-wide metabolic fitness landscape of the bacterium Escherichia coli Deformability is quantified by the noncommutativity of epistatic interactions, which we experimentally demonstrate in mutant strains on the path to an evolutionary innovation. Our analysis shows that the deformation of fitness landscapes by metabolic mutations rarely affects evolutionary trajectories in the short range. However, mutations with large environmental effects produce long-range landscape deformations in distant regions of the genotype space that affect the fitness of later descendants. Our results therefore suggest that, even in situations in which mutations have strong environmental effects, fitness landscapes may retain their power to forecast evolution over small mutational distances despite the potential attenuation of that power over longer evolutionary trajectories. Our methods and results provide an avenue for integrating adaptive and eco-evolutionary dynamics with complex genetics and genomics.
Keywords: eco-evolutionary feedbacks; ecologically mediated gene interactions; fitness landscapes; gene × environment × gene interactions; noncommutative epistasis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Lewontin RC. The organism as the subject and object of evolution. Scientia. 1983;77:65.
-
- John Odling-Smee F, Laland KN, Feldman MW. Niche Construction: The Neglected Process in Evolution (MPB-37) Princeton Univ Press; Princeton: 2013.
-
- Laland K, et al. Does evolutionary theory need a rethink? Nature. 2014;514:161–164. - PubMed
-
- Hendry AP. Eco-Evolutionary Dynamics. Princeton Univ Press; Princeton: 2016.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
