

Gap Junctions and NCA Cation Channels Are Critical for Developmentally Timed Sleep and Arousal in Caenorhabditis elegans
- PMID: 30323068
- PMCID: PMC6283151
- DOI: 10.1534/genetics.118.301551
Gap Junctions and NCA Cation Channels Are Critical for Developmentally Timed Sleep and Arousal in Caenorhabditis elegans
Abstract
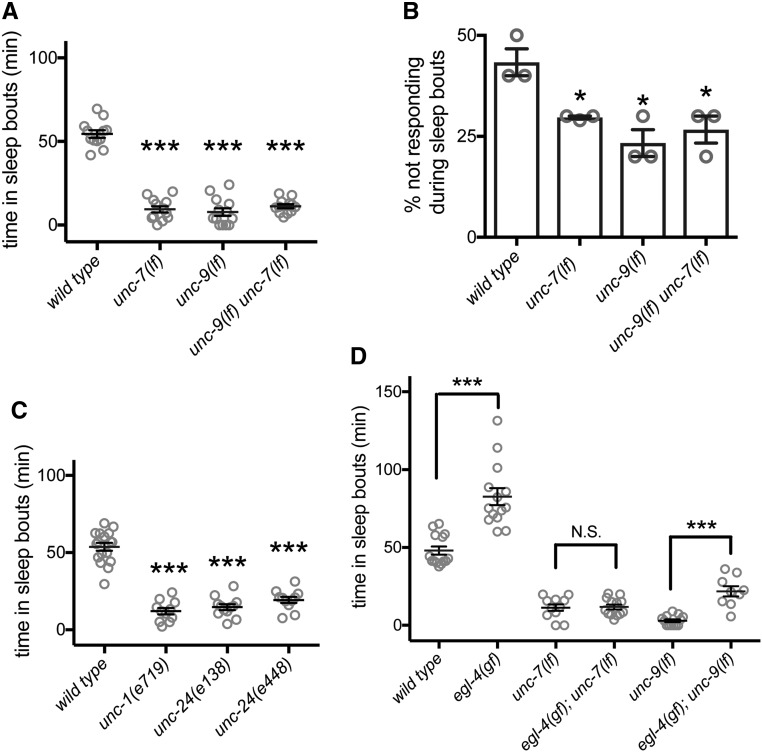
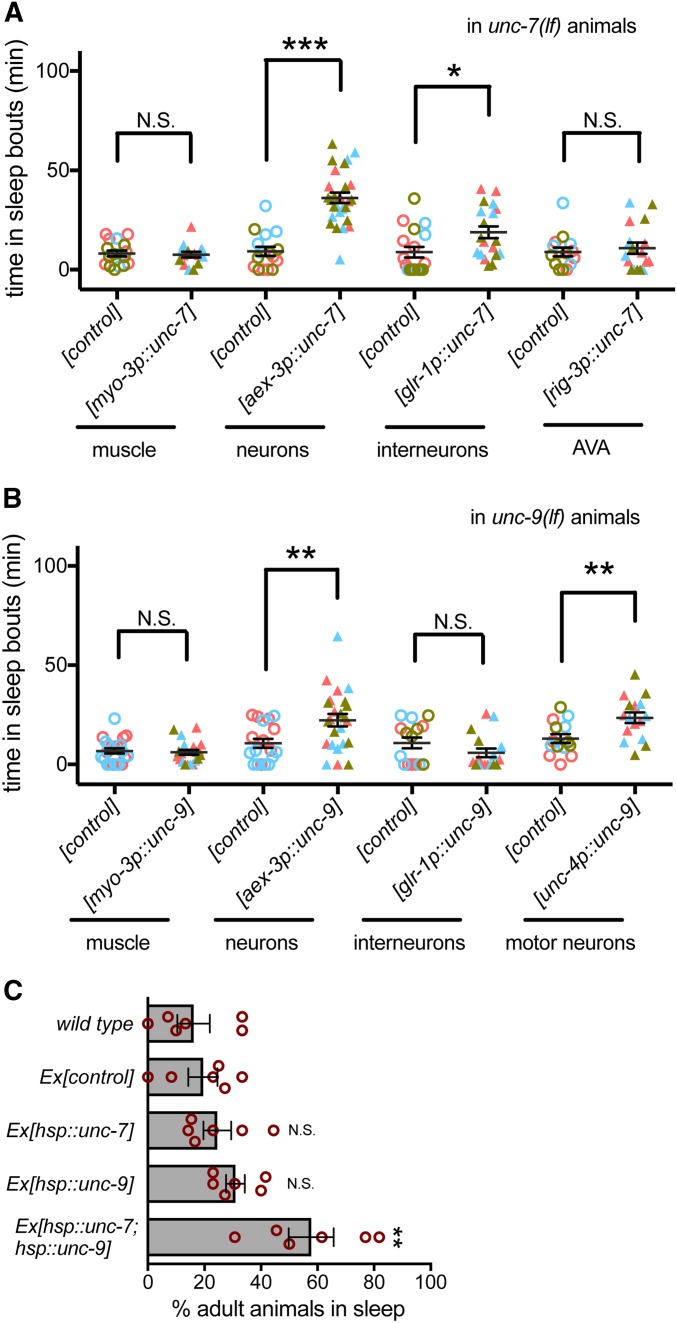
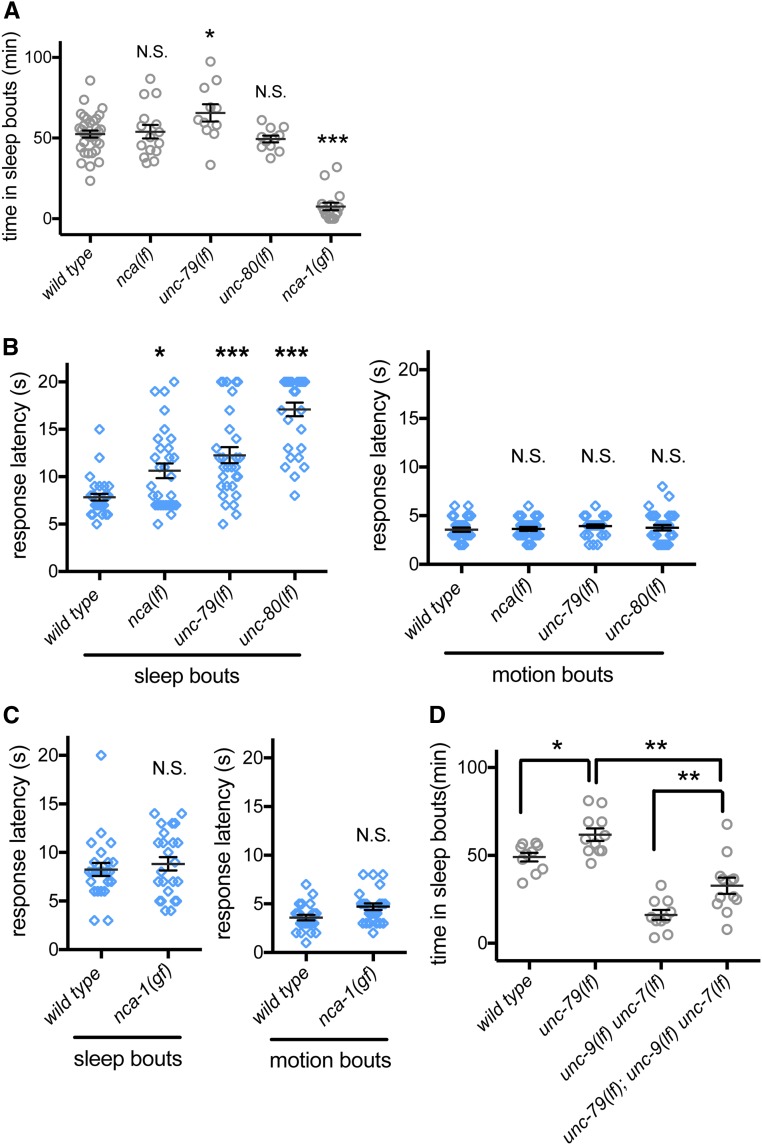
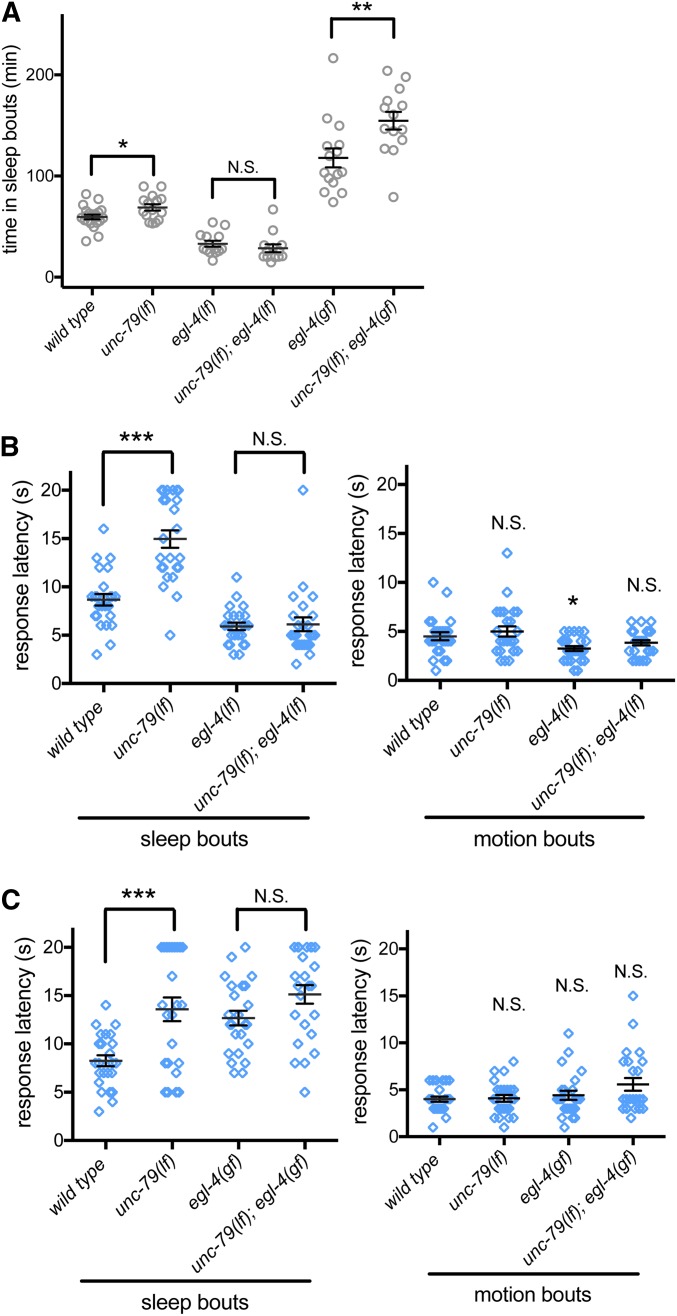
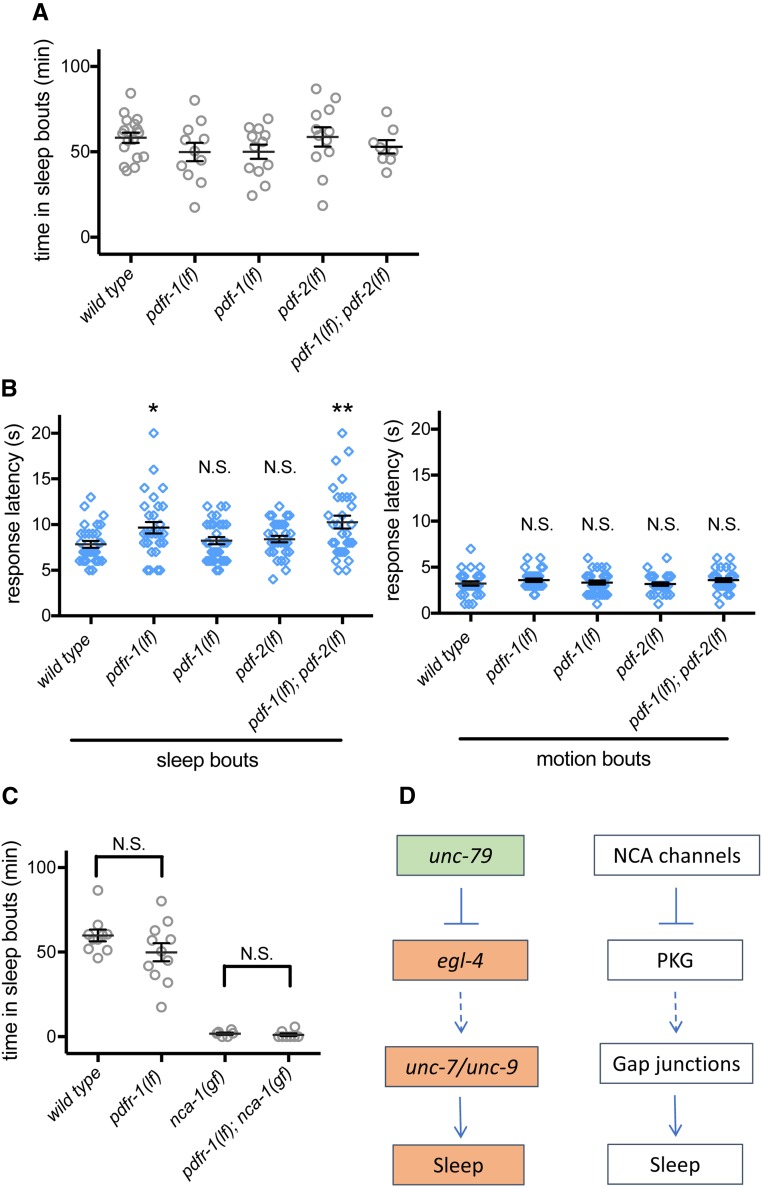
An essential characteristic of sleep is heightened arousal threshold, with decreased behavioral response to external stimuli. The molecular and cellular mechanisms underlying arousal threshold changes during sleep are not fully understood. We report that loss of UNC-7 or UNC-9 innexin function dramatically reduced sleep and decreased arousal threshold during developmentally timed sleep in Caenorhabditiselegans UNC-7 function was required in premotor interneurons and UNC-9 function was required in motor neurons in this paradigm. Simultaneous transient overexpression of UNC-7 and UNC-9 was sufficient to induce anachronistic sleep in adult animals. Moreover, loss of UNC-7 or UNC-9 suppressed the increased sleep of EGL-4 gain-of-function animals, which have increased cyclic-GMP-dependent protein kinase activity. These results suggest C. elegans gap junctions may act downstream of previously identified sleep regulators. In other paradigms, the NCA cation channels act upstream of gap junctions. Consistent with this, diminished NCA channel activity in C. elegans robustly increased arousal thresholds during sleep bouts in L4-to-adult developmentally timed sleep. Total time in sleep bouts was only modestly increased in animals lacking NCA channel auxiliary subunit UNC-79, whereas increased channel activity dramatically decreased sleep. Loss of EGL-4 or innexin proteins suppressed UNC-79 loss-of-function sleep and arousal defects. In Drosophila, the ion channel narrow abdomen, an ortholog of the C. elegans NCA channels, drive the pigment dispersing factor (PDF) neuropeptide release, regulating circadian behavior. However, in C. elegans, we found that loss of the PDF receptor PDFR-1 did not suppress gain-of-function sleep defects, suggesting an alternative downstream pathway. This study emphasizes the conservation and importance of neuronal activity modulation during sleep, and unequivocally demonstrates that gap junction function is critical for normal sleep.
Keywords: Caenorhabditis elegans sleep; NCA channel; cGMP-dependent kinase; gap junction.
Copyright © 2018 by the Genetics Society of America.
Figures
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
