

The Wheat MYB Transcription Factor TaMYB31 Is Involved in Drought Stress Responses in Arabidopsis
- PMID: 30323824
- PMCID: PMC6172359
- DOI: 10.3389/fpls.2018.01426
The Wheat MYB Transcription Factor TaMYB31 Is Involved in Drought Stress Responses in Arabidopsis
Abstract
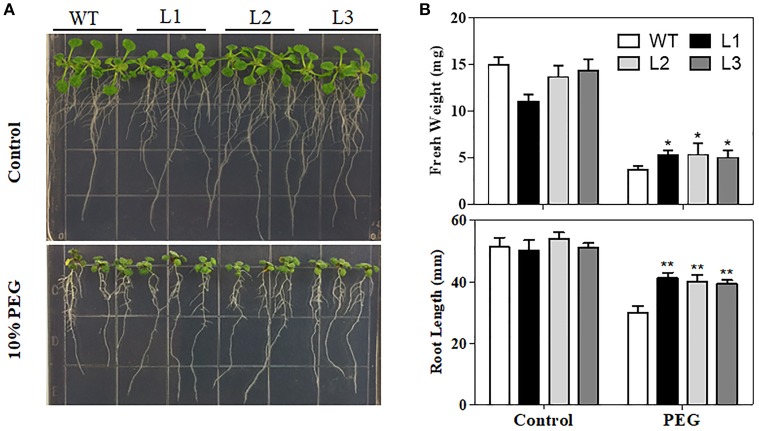
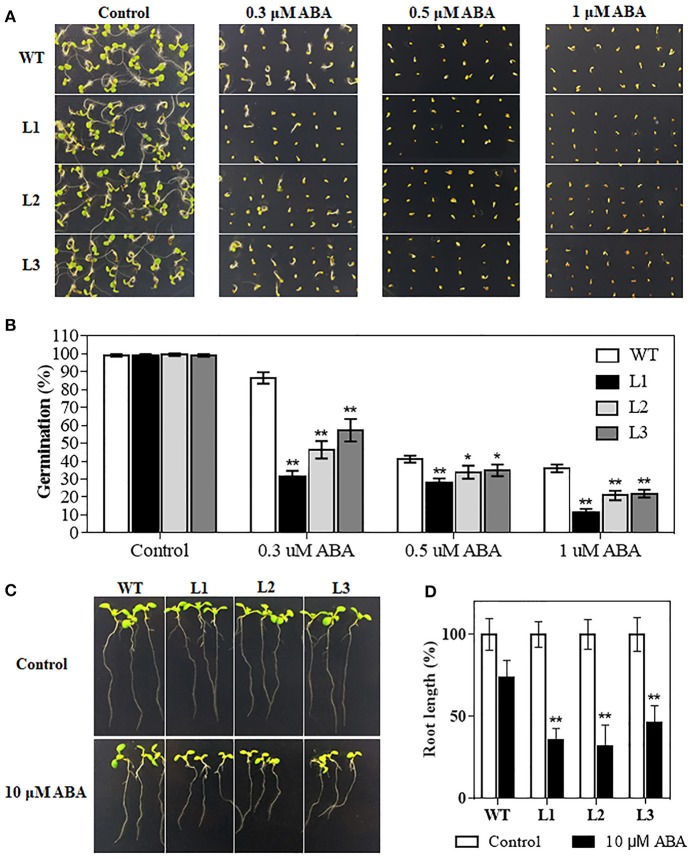
Drought is one of the major environmental stresses limiting crop growth and production. MYB family transcription factors play crucial roles in response to abiotic stresses. Previous studies found that TaMYB31 is transcriptionally induced by drought stress. However, the biological functions of TaMYB31 in drought stress responses remained unknown. In this study, three TaMYB31 homoeologous genes from hexaploid wheat, designated TaMYB31-A, TaMYB31-B, and TaMYB31-D, were cloned and characterized. Expression analysis showed that TaMYB31 genes have different tissue expression patterns, and TaMYB31-B has relatively high expression levels in most tested tissues. All the three homoeologs were up-regulated by polyethylene glycol (PEG) 6000 and abscisic acid (ABA) treatments. Subcellular localization analyses revealed that TaMYB31 is localized to the nucleus. Ectopic expression of the TaMYB31-B gene in Arabidopsis affected plants growth and enhanced drought tolerance. In addition, seed germination and seedling root growth of TaMYB31-B transgenic plants were more sensitive to exogenous ABA treatment compared to wild type control. RNA-seq analysis indicated that TaMYB31 functions through up-regulation of wax biosynthesis genes and drought-responsive genes. These results provide evidence that TaMYB31 acts as a positive regulator of drought resistance, and justify its potential application in genetic modification of crop drought tolerance.
Keywords: Arabidopsis; MYB; RNA-seq; drought; transgenic; wheat.
Figures






References
-
- Aharoni A., Dixit S., Jetter R., Thoenes E., van Arkel G., Pereira A. (2004). The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis. Plant Cell 16, 2463–2480. 10.1105/tpc.104.022897 - DOI - PMC - PubMed
-
- Bi H., Shi J., Kovalchuk N., Luang S., Bazanova N., Chirkova L., et al. . (2018). Overexpression of the TaSHN1 transcription factor in bread wheat leads to leaf surface modifications, improved drought tolerance and no yield penalty under controlled growth conditions. Plant Cell Environ. [Epub ahead of print]. 10.1111/pce.13339 - DOI - PubMed
LinkOut - more resources
Full Text Sources