

Cellulose synthase-like D1 controls organ size in maize
- PMID: 30326832
- PMCID: PMC6192064
- DOI: 10.1186/s12870-018-1453-8
Cellulose synthase-like D1 controls organ size in maize
Abstract
Background: Plant architecture is a critical factor that affects planting density and, consequently, grain yield in maize. The genes or loci that determine organ size are the key regulators of plant architecture. Thus, understanding the genetic and molecular mechanisms of organ size will inform the use of a molecular manipulation approach to improve maize plant architecture and grain yield.
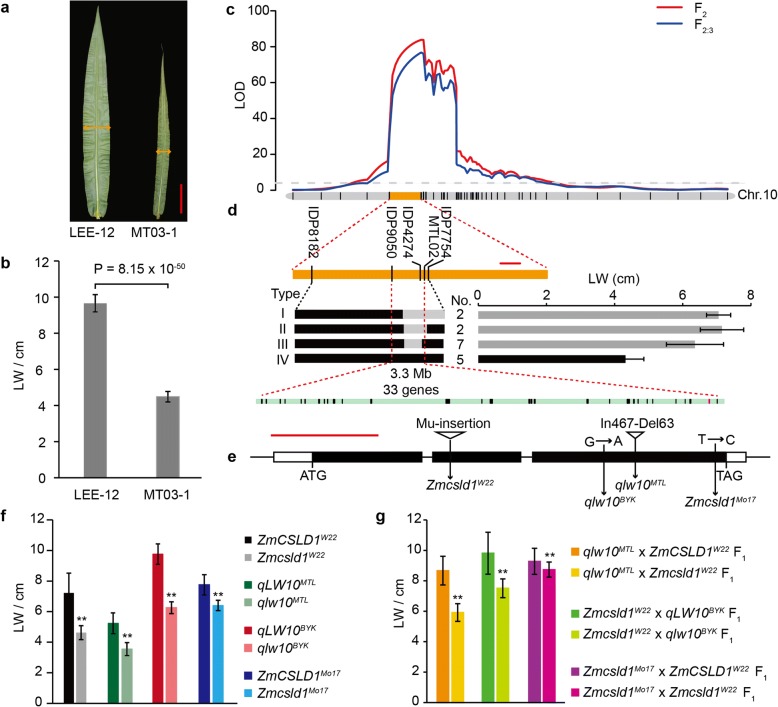
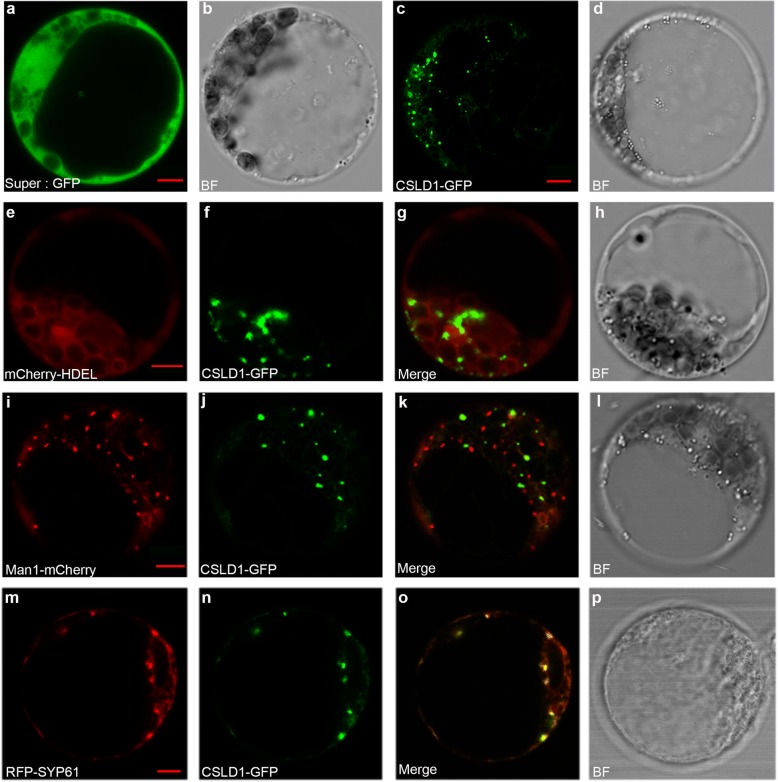
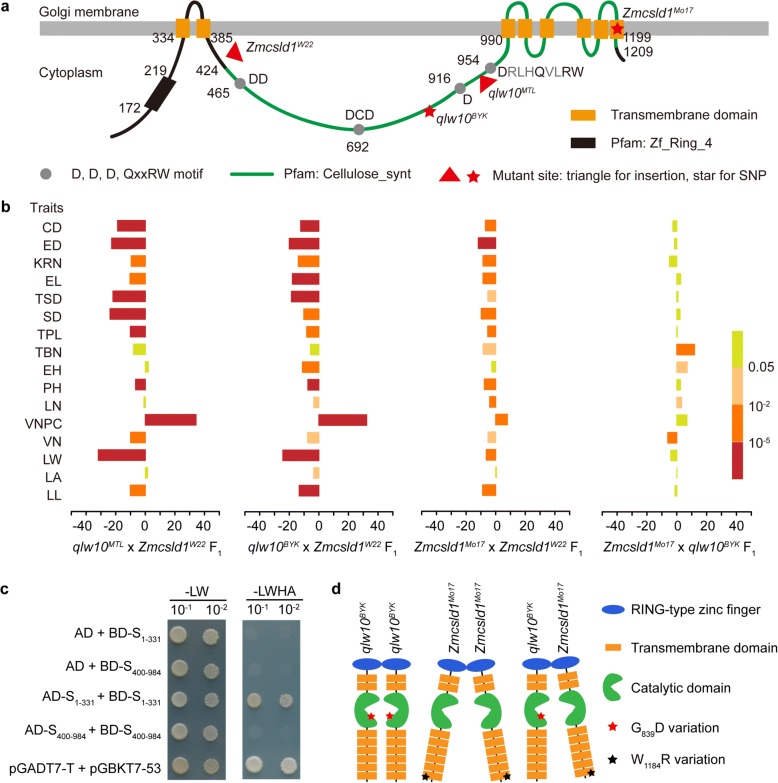
Results: A total of 18 unique quantitative trait loci (QTLs) were identified for 11 agronomic traits in the F2 and F2:3 segregating populations derived from a cross between a double haploid line with a small plant architecture (MT03-1) and an inbred line with a large plant architecture (LEE-12). Subsequently, we showed that one QTL, qLW10, for multiple agronomic traits that relate to plant organ size reflects allelic variation in ZmCSLD1, which encodes a cellulose synthase-like D protein. ZmCSLD1 was localized to the trans-Golgi and was highly expressed in the rapidly growing regions. The loss of ZmCSLD1 function decreased cell division, which resulted in smaller organs with fewer cell numbers and, in turn, pleiotropic effects on multiple agronomic traits. In addition, intragenic complementation was investigated for two Zmcsld1 alleles with nonsynonymous SNPs in different functional domains, and the mechanism of this complementation was determined to be through homodimeric interactions.
Conclusions: Through positional cloning by using two populations and allelism tests, qLW10 for organ size was resolved to be a cellulose synthase-like D family gene, ZmCSLD1. ZmCSLD1 has pleiotropic effects on multiple agronomic traits that alter plant organ size by changing the process of cell division. These findings provide new insight into the regulatory mechanism that underlies plant organ development.
Keywords: Organ size; Pleiotropic effects and intragenic complementation; QTL; ZmCSLD1.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures



References
-
- Duvick DN. The contribution of breeding to yield advances in maize (Zea mays L.) Adv Agron. 2005;86:83–145. doi: 10.1016/S0065-2113(05)86002-X. - DOI
-
- Fischer K, Edmeades G, Johnson E. Recurrent selection for reduced tassel branch number and reduced leaf area density above the ear in tropical maize populations. Crop Sci. 1987;27:1150–1156. doi: 10.2135/cropsci1987.0011183X002700060013x. - DOI
-
- Johnson E, Fischer K, Edmeades G, Palmer A. Recurrent selection for reduced plant height in lowland tropical maize. Crop Sci. 1986;26:253–260. doi: 10.2135/cropsci1986.0011183X002600020008x. - DOI
-
- Schuetz SH, Mock JJ. Genetics of tassel branch number in maize and its implications for a selection program for small tassel size. Theor Appl Genet. 1978;53:265–271. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
