

Peripherally derived macrophages modulate microglial function to reduce inflammation after CNS injury
- PMID: 30332405
- PMCID: PMC6205650
- DOI: 10.1371/journal.pbio.2005264
Peripherally derived macrophages modulate microglial function to reduce inflammation after CNS injury
Abstract
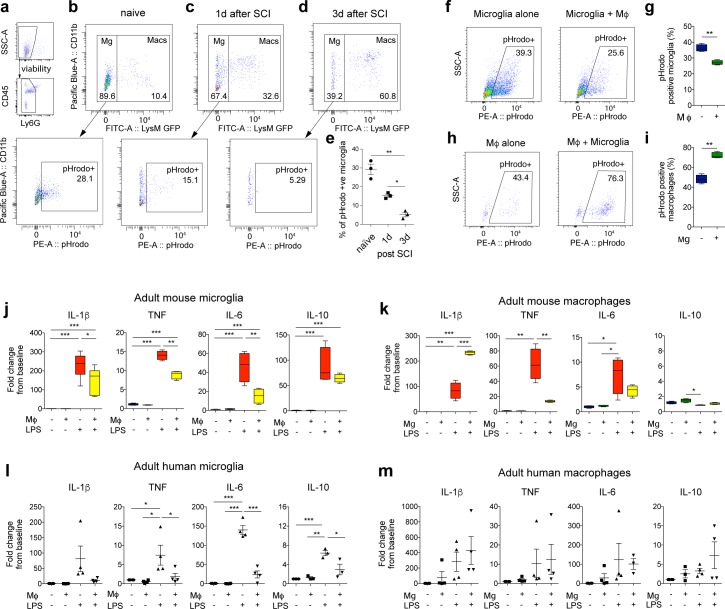
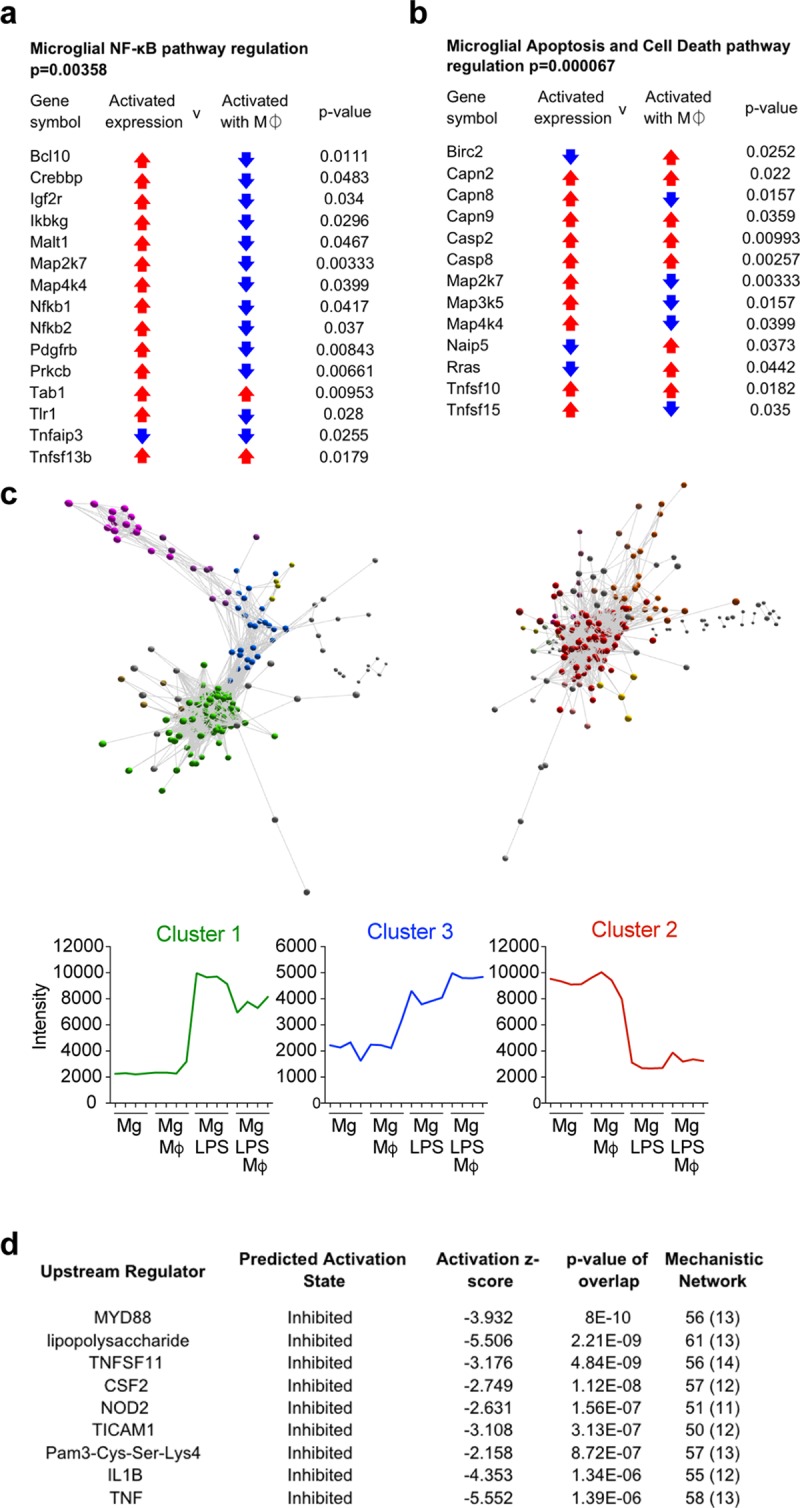
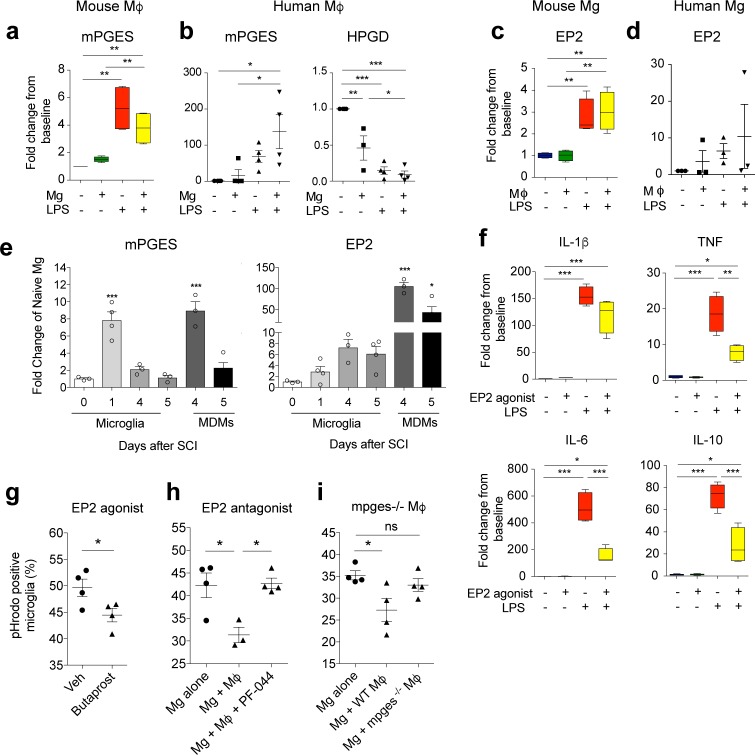
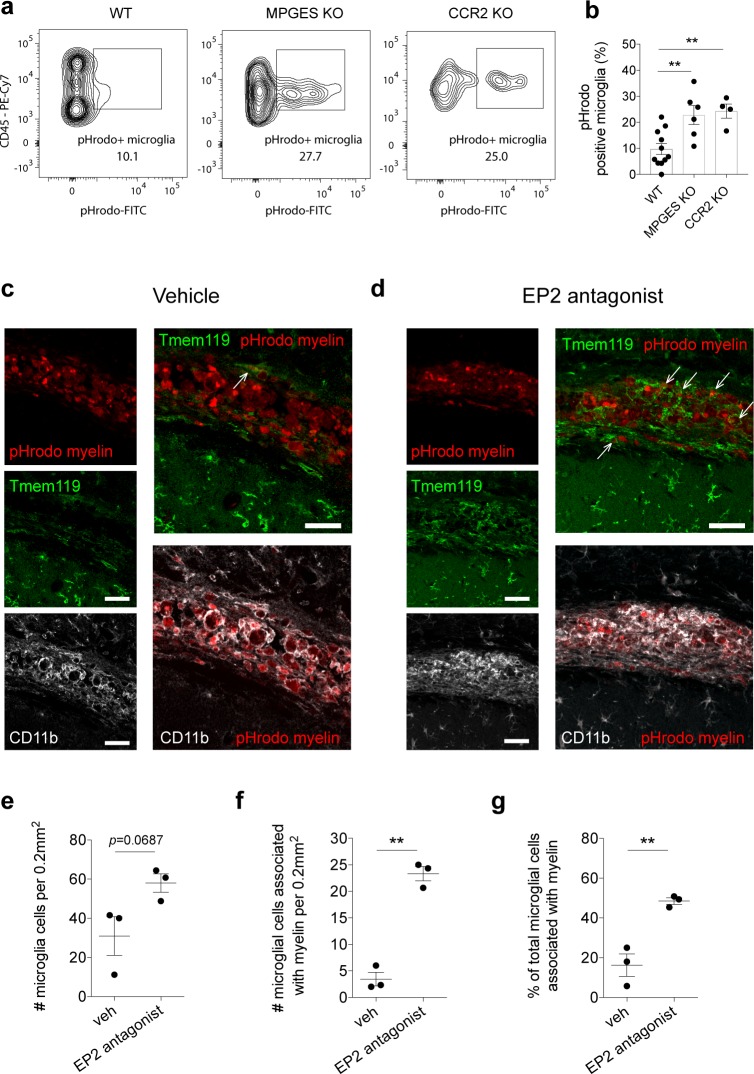
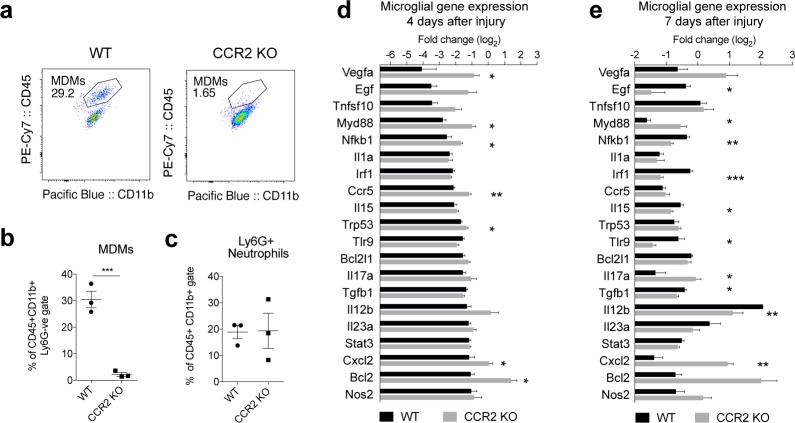
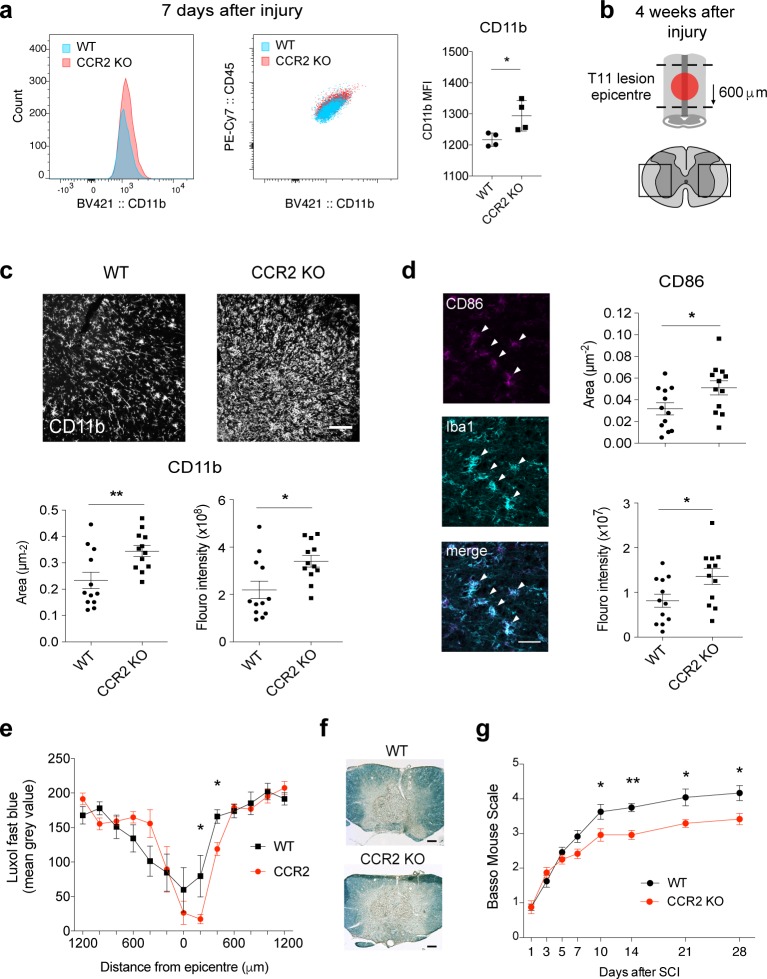
Infiltrating monocyte-derived macrophages (MDMs) and resident microglia dominate central nervous system (CNS) injury sites. Differential roles for these cell populations after injury are beginning to be uncovered. Here, we show evidence that MDMs and microglia directly communicate with one another and differentially modulate each other's functions. Importantly, microglia-mediated phagocytosis and inflammation are suppressed by infiltrating macrophages. In the context of spinal cord injury (SCI), preventing such communication increases microglial activation and worsens functional recovery. We suggest that macrophages entering the CNS provide a regulatory mechanism that controls acute and long-term microglia-mediated inflammation, which may drive damage in a variety of CNS conditions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
