

Rosiglitazone ameliorates palmitic acid-induced cytotoxicity in TM4 Sertoli cells
- PMID: 30333041
- PMCID: PMC6192158
- DOI: 10.1186/s12958-018-0416-0
Rosiglitazone ameliorates palmitic acid-induced cytotoxicity in TM4 Sertoli cells
Abstract
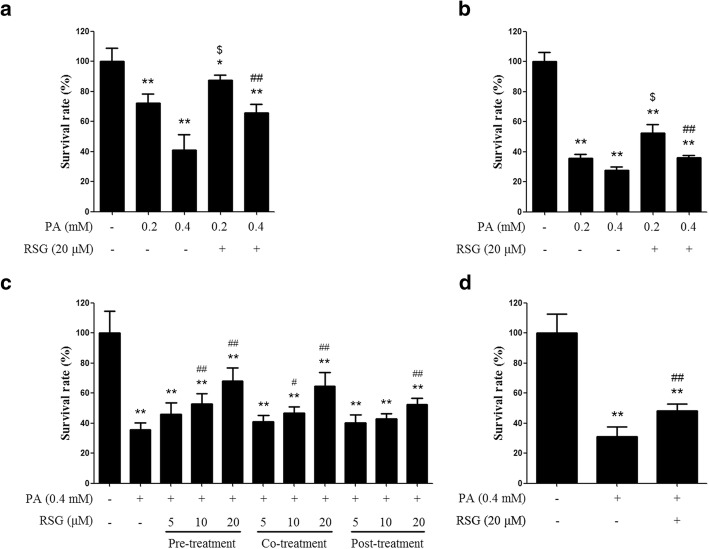
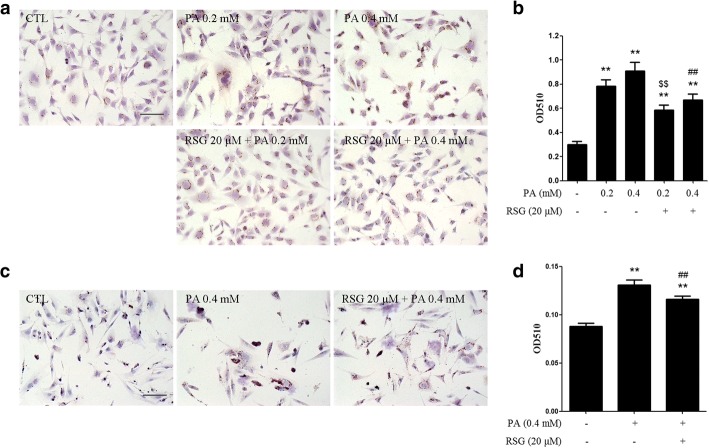
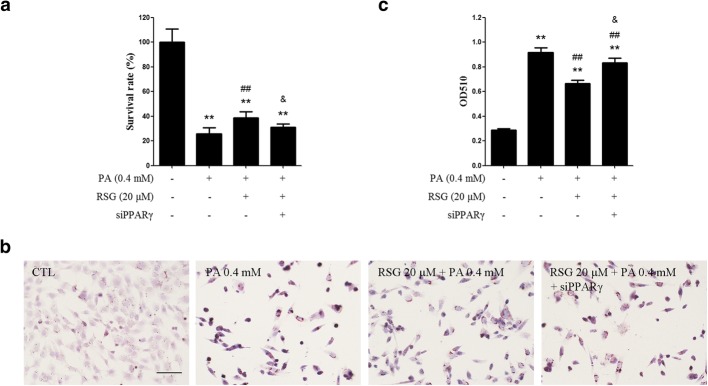
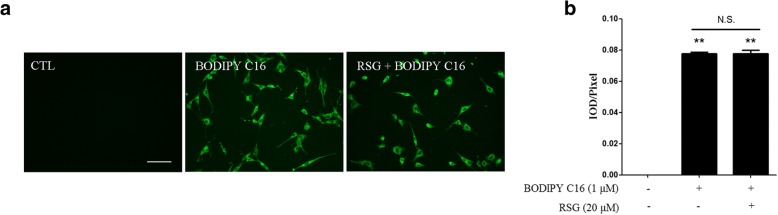
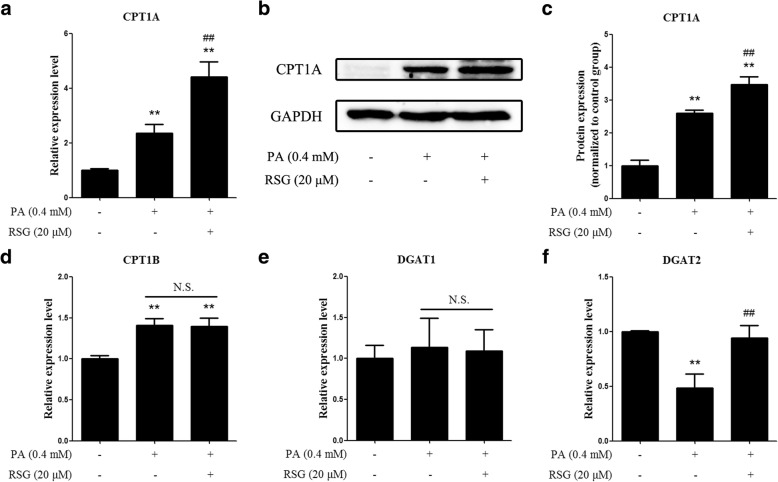
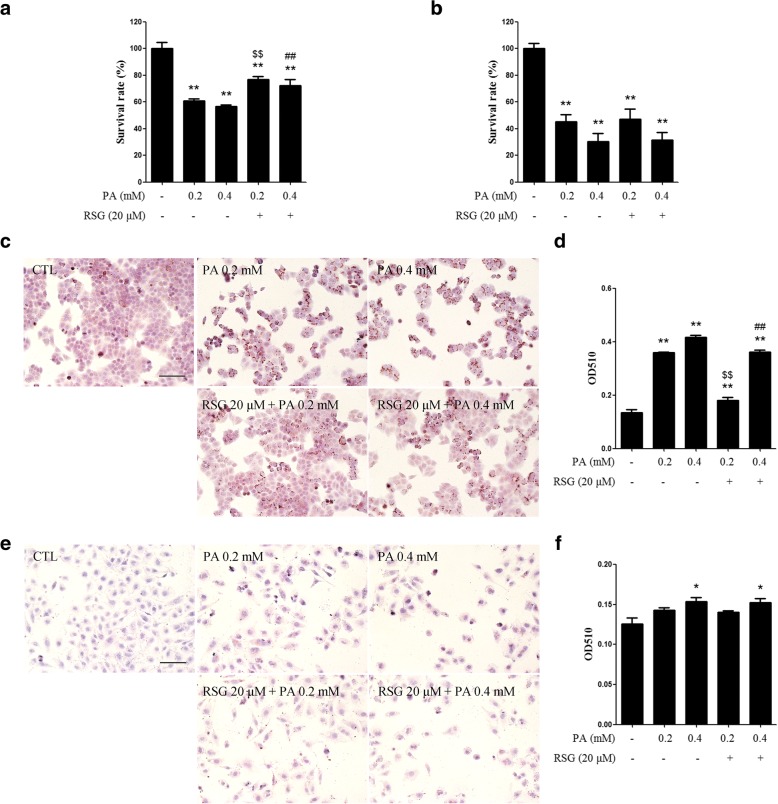
The Sertoli cell is the only somatic cell within the seminiferous tubules, and is vital for testis development and spermatogenesis. Rosiglitazone (RSG) is a member of the thiazolidinedione family and is a peroxisome proliferator-activated receptor-γ (PPARγ) agonist. It has been reported that RSG protects various types of cells from fatty acid-induced damage. However, whether RSG serves a protective role in Sertoli cells against palmitic acid (PA)-induced toxicity remains to be elucidated. Therefore, the aim of the present study was to investigate the effect of RSG on PA-induced cytotoxicity in Sertoli cells. MTT assay and Oil Red O staining revealed that RSG ameliorated the PA-induced decrease in TM4 cell viability, which was accompanied by an alleviation of PA-induced lipid accumulation in cells. In primary mouse Sertoli cells, RSG also showed similar protective effects against PA-induced lipotoxicity. Knockdown of PPARγ verified that RSG exerted its protective role in TM4 cells through a PPARγ-dependent pathway. To evaluate the mechanism underlying the protective role of RSG on PA-induced lipotoxicity, the present study analyzed the effects of RSG on PA uptake, and the expression of genes associated with both fatty acid oxidation and triglyceride synthesis. The results demonstrated that although RSG did not affect the endocytosis of PA, it significantly elevated the expression of carnitine palmitoyltransferase (CPT)-1A, a key enzyme involved in fatty acid oxidation, which indicated that the protective effect of RSG may have an important role in fatty acid oxidation. On the other hand, the expression of CPT1B was not affected by RSG. Moreover, the expression levels of diacylglycerol O-acyltransferase (DGAT)-1 and DGAT2, both of which encode enzymes catalyzing the synthesis of triglycerides, were not suppressed by RSG. The results indicated that RSG reduced PA-induced lipid accumulation by promoting fatty acid oxidation mediated by CPT1A. The effect of RSG in protecting cells from lipotoxicity was also found to be specific to Sertoli cells and hepatocytes, and not to other cell types that do not store excess lipid in large quantities, such as human umbilical vein endothelial cells. These findings provide insights into the cytoprotective effects of RSG on Sertoli cells and suggest that PPARγ activation may be a useful therapeutic method for the treatment of Sertoli cell dysfunction caused by dyslipidemia.
Keywords: Cytotoxicity; Palmitic acid; Rosiglitazone; Sertoli cells.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Nery SF, Vieira MA, Dela Cruz C, Lobach VN, Del Puerto HL, Torres PB, Rocha AL, Reis AB, Reis FM. Seminal plasma concentrations of anti-Mullerian hormone and inhibin B predict motile sperm recovery from cryopreserved semen in asthenozoospermic men: a prospective cohort study. Andrology. 2014;2(6):918–923. doi: 10.1111/andr.278. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
