Oxidative stress in sperm affects the epigenetic reprogramming in early embryonic development
- PMID: 30333056
- PMCID: PMC6192351
- DOI: 10.1186/s13072-018-0224-y
Oxidative stress in sperm affects the epigenetic reprogramming in early embryonic development
Abstract
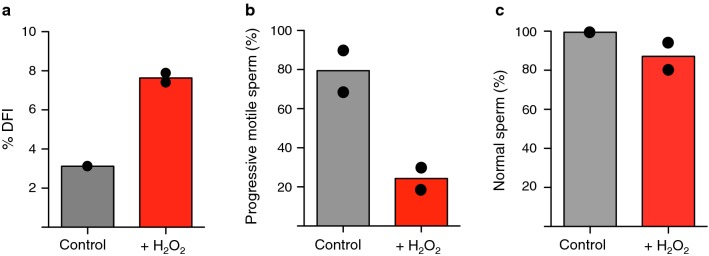
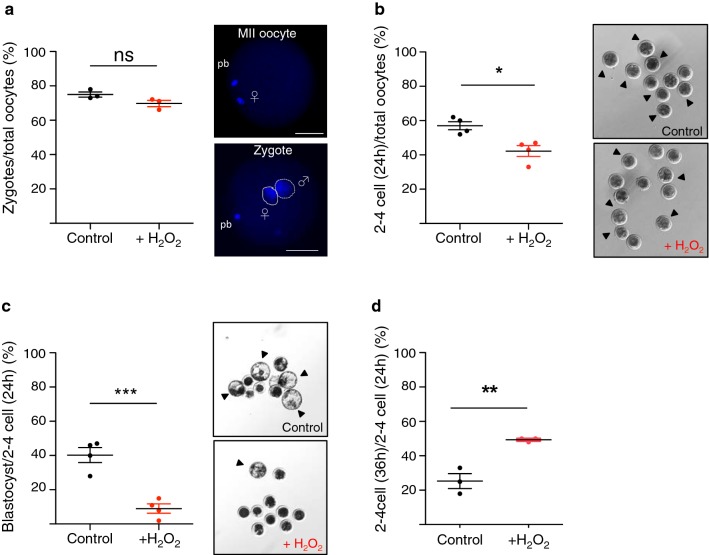
Background: Reactive oxygen species (ROS)-induced oxidative stress is well known to play a major role in male infertility. Sperm are sensitive to ROS damaging effects because as male germ cells form mature sperm they progressively lose the ability to repair DNA damage. However, how oxidative DNA lesions in sperm affect early embryonic development remains elusive.
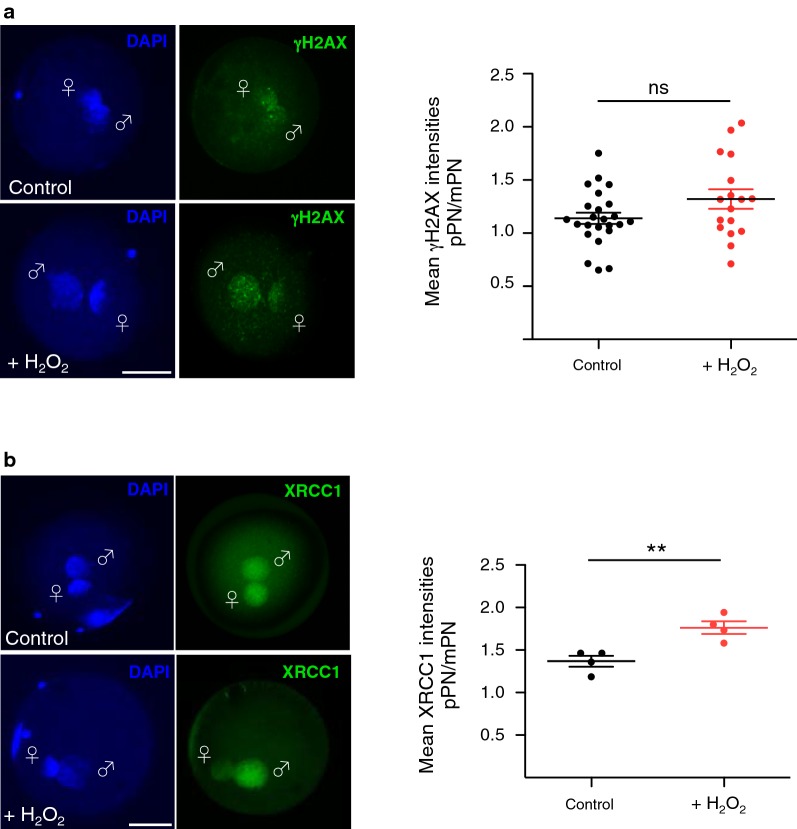
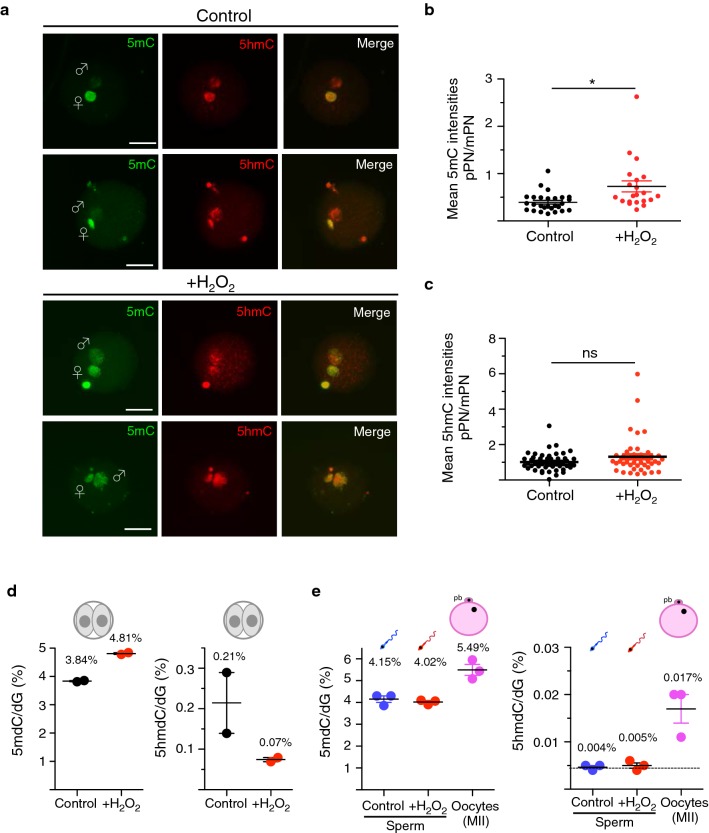
Results: Using cattle as model, we show that fertilization using sperm exposed to oxidative stress caused a major developmental arrest at the time of embryonic genome activation. The levels of DNA damage response did not directly correlate with the degree of developmental defects. The early cellular response for DNA damage, γH2AX, is already present at high levels in zygotes that progress normally in development and did not significantly increase at the paternal genome containing oxidative DNA lesions. Moreover, XRCC1, a factor implicated in the last step of base excision repair (BER) pathway, was recruited to the damaged paternal genome, indicating that the maternal BER machinery can repair these DNA lesions induced in sperm. Remarkably, the paternal genome with oxidative DNA lesions showed an impairment of zygotic active DNA demethylation, a process that previous studies linked to BER. Quantitative immunofluorescence analysis and ultrasensitive LC-MS-based measurements revealed that oxidative DNA lesions in sperm impair active DNA demethylation at paternal pronuclei, without affecting 5-hydroxymethylcytosine (5hmC), a 5-methylcytosine modification that has been implicated in paternal active DNA demethylation in mouse zygotes. Thus, other 5hmC-independent processes are implicated in active DNA demethylation in bovine embryos. The recruitment of XRCC1 to damaged paternal pronuclei indicates that oxidative DNA lesions drive BER to repair DNA at the expense of DNA demethylation. Finally, this study highlighted striking differences in DNA methylation dynamics between bovine and mouse zygotes that will facilitate the understanding of the dynamics of DNA methylation in early development.
Conclusions: The data demonstrate that oxidative stress in sperm has an impact not only on DNA integrity but also on the dynamics of epigenetic reprogramming, which may harm the paternal genetic and epigenetic contribution to the developing embryo and affect embryo development and embryo quality.
Keywords: BER; DNA methylation; Epigenetic reprogramming; Oxidative stress.
Figures