

Acute Infection and Subsequent Subclinical Reactivation of Herpes Simplex Virus 2 after Vaginal Inoculation of Rhesus Macaques
- PMID: 30333177
- PMCID: PMC6321901
- DOI: 10.1128/JVI.01574-18
Acute Infection and Subsequent Subclinical Reactivation of Herpes Simplex Virus 2 after Vaginal Inoculation of Rhesus Macaques
Abstract
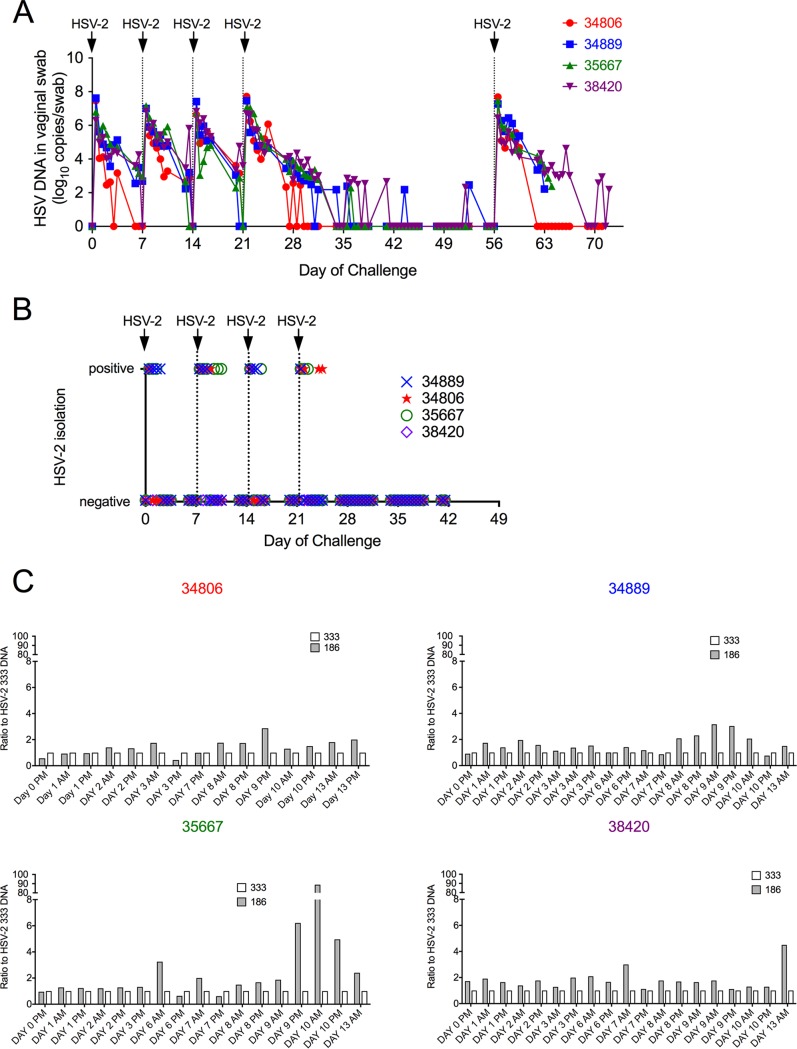
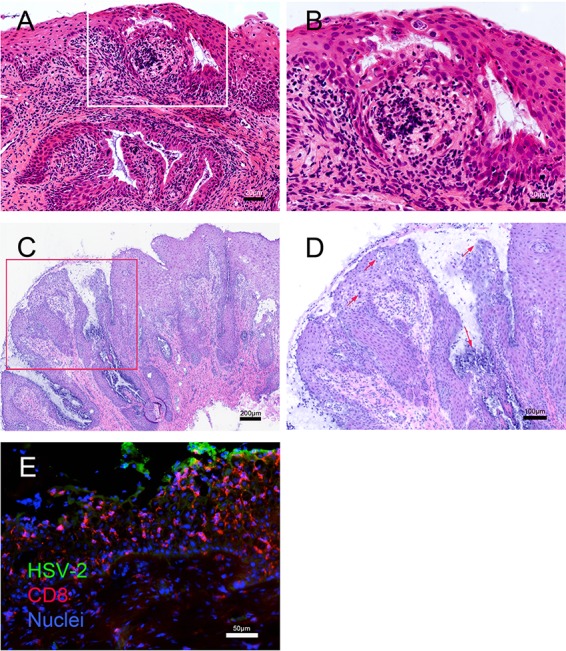
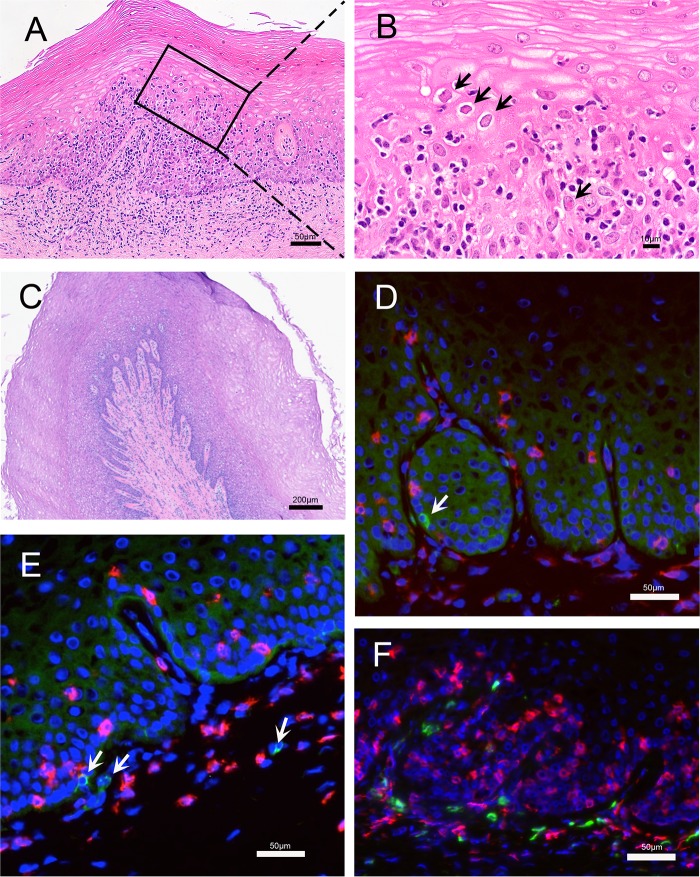
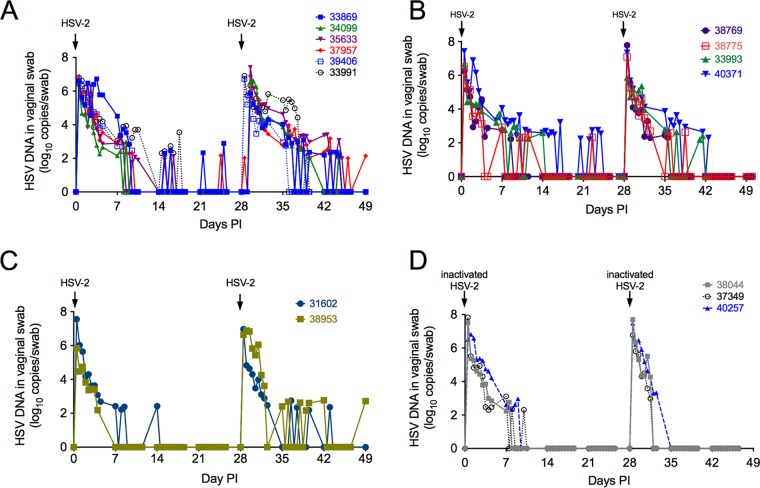
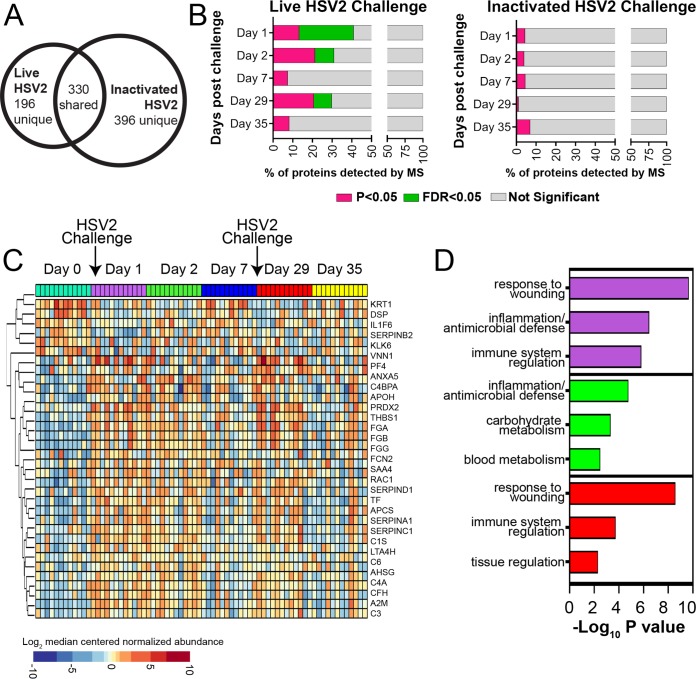
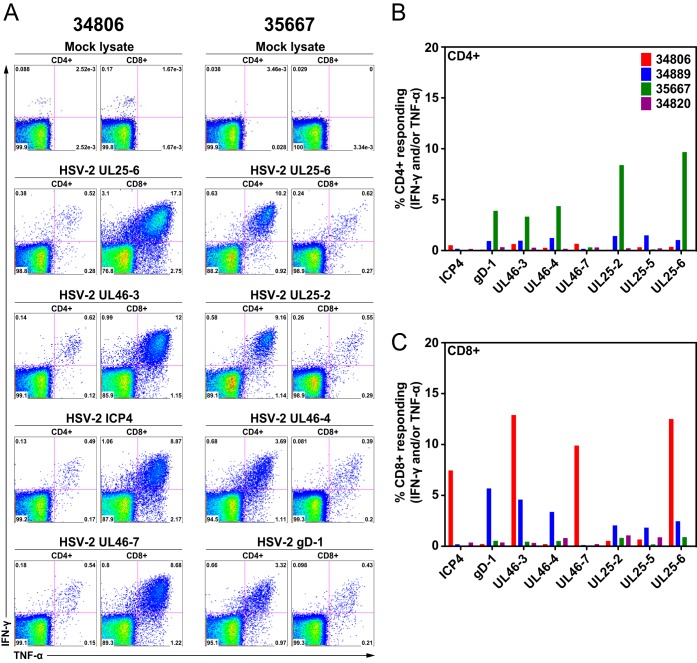
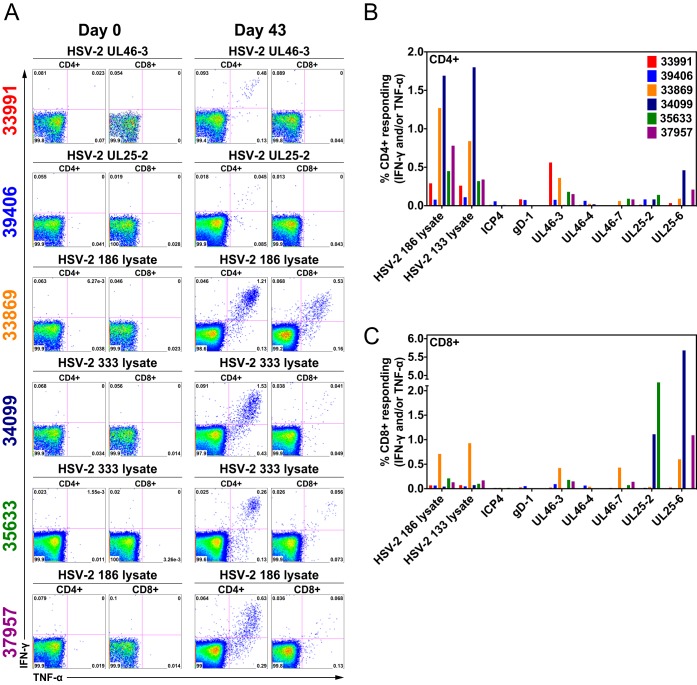
Herpes simplex virus 2 (HSV-2) is a common sexually transmitted infection with a highly variable clinical course. Many infections quickly become subclinical, with episodes of spontaneous virus reactivation. To study host-HSV-2 interactions, an animal model of subclinical HSV-2 infection is needed. In an effort to develop a relevant model, rhesus macaques (RM) were inoculated intravaginally with two or three HSV-2 strains (186, 333, and/or G) at a total dose of 1 × 107 PFU of HSV-2 per animal. Infectious HSV-2 and HSV-2 DNA were consistently shed in vaginal swabs for the first 7 to 14 days after each inoculation. Proteins associated with wound healing, innate immunity, and inflammation were significantly increased in cervical secretions immediately after HSV-2 inoculation. There was histologic evidence of acute herpesvirus pathology, including acantholysis in the squamous epithelium and ballooning degeneration of and intranuclear inclusion bodies in epithelial cells, with HSV antigen in mucosal epithelial cells and keratinocytes. Further, an intense inflammatory infiltrate was found in the cervix and vulva. Evidence of latent infection and reactivation was demonstrated by the detection of spontaneous HSV-2 shedding post-acute inoculation (102 to 103 DNA copies/swab) in 80% of RM. Further, HSV-2 DNA was detected in ganglia in most necropsied animals. HSV-2-specifc T-cell responses were detected in all animals, although antibodies to HSV-2 were detected in only 30% of the animals. Thus, HSV-2 infection of RM recapitulates many of the key features of subclinical HSV-2 infection in women but seems to be more limited, as virus shedding was undetectable more than 40 days after the last virus inoculation.IMPORTANCE Herpes simplex virus 2 (HSV-2) infects nearly 500 million persons globally, with an estimated 21 million incident cases each year, making it one of the most common sexually transmitted infections (STIs). HSV-2 is associated with increased human immunodeficiency virus type 1 (HIV-1) acquisition, and this risk does not decline with the use of antiherpes drugs. As initial acquisition of both HIV and HSV-2 infections is subclinical, study of the initial molecular interactions of the two agents requires an animal model. We found that HSV-2 can infect RM after vaginal inoculation, establish latency in the nervous system, and spontaneously reactivate; these features mimic some of the key features of HSV-2 infection in women. RM may provide an animal model to develop strategies to prevent HSV-2 acquisition and reactivation.
Keywords: T-cell responses; cervix; female reproductive tract; herpes simplex virus; proteomics; subclinical infection; vaginal swabs.
Copyright © 2019 Lo et al.
Figures
References
-
- Gottlieb MS, Schanker HM, Fan PT. 1981. Pneumocystis pneumonia—Los Angeles. MMWR Morb Mortal Wkly Rep 30:250–252. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
