

Activation of Serotonin 5-HT7 Receptors Modulates Hippocampal Synaptic Plasticity by Stimulation of Adenylate Cyclases and Rescues Learning and Behavior in a Mouse Model of Fragile X Syndrome
- PMID: 30333723
- PMCID: PMC6176069
- DOI: 10.3389/fnmol.2018.00353
Activation of Serotonin 5-HT7 Receptors Modulates Hippocampal Synaptic Plasticity by Stimulation of Adenylate Cyclases and Rescues Learning and Behavior in a Mouse Model of Fragile X Syndrome
Abstract
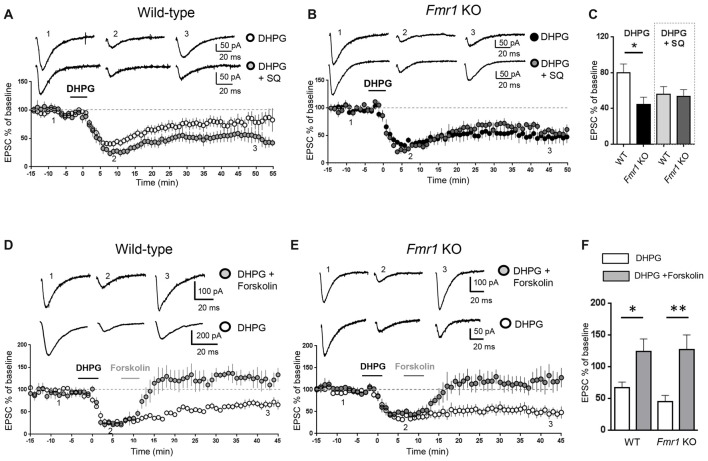
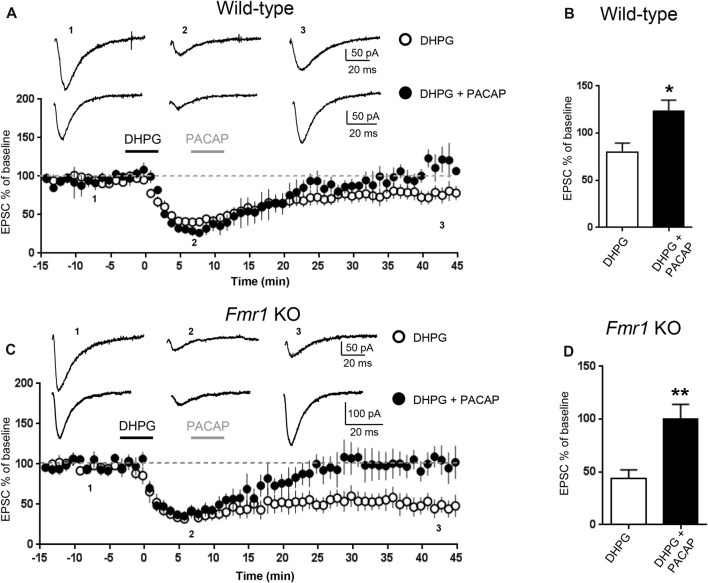
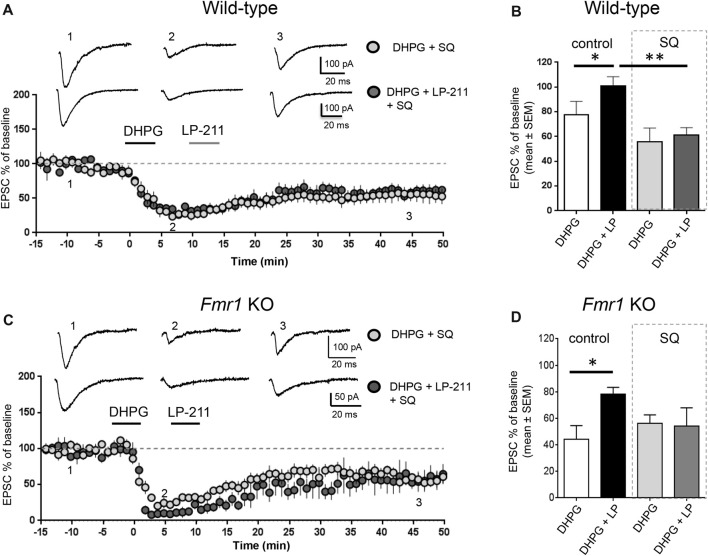
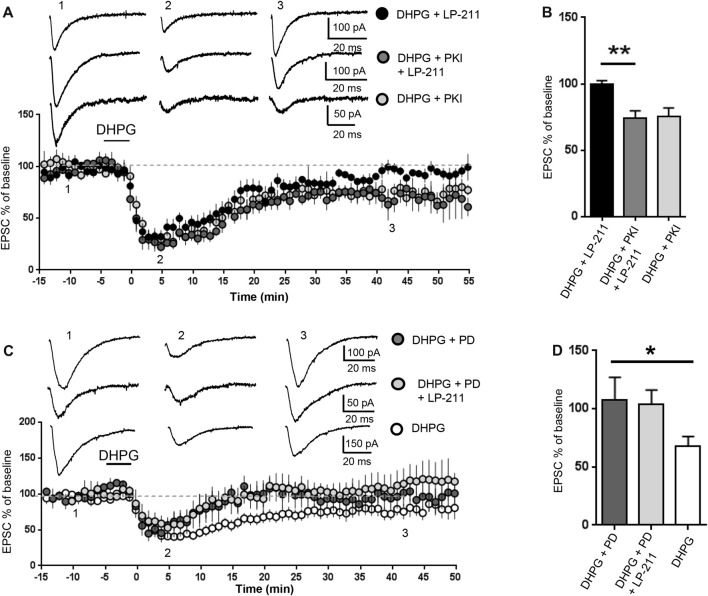
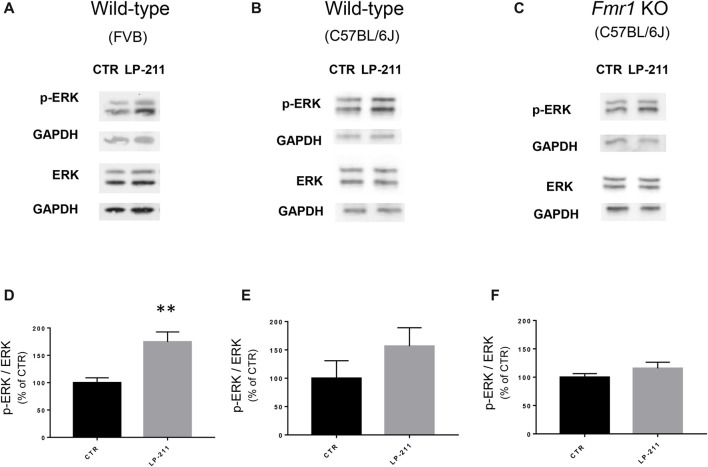
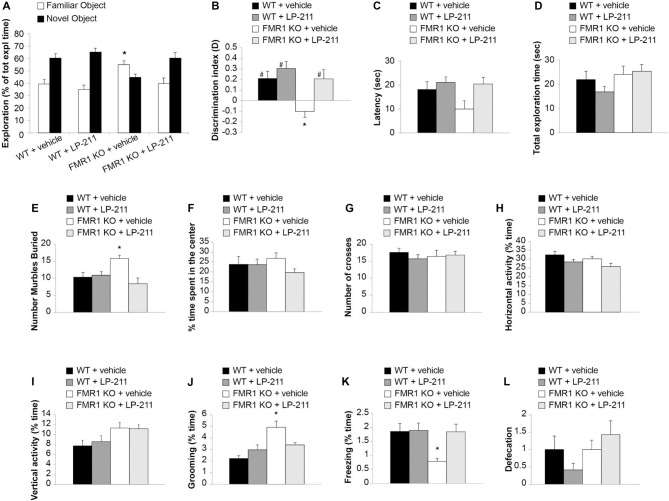
We have previously demonstrated that activation of serotonin 5-HT7 receptors (5-HT7R) reverses metabotropic glutamate receptor-mediated long term depression (mGluR-LTD) in the hippocampus of wild-type (WT) and Fmr1 Knockout (KO) mice, a model of Fragile X Syndrome (FXS) in which mGluR-LTD is abnormally enhanced. Here, we have investigated intracellular mechanisms underlying the effect of 5-HT7R activation using patch clamp on hippocampal slices. Furthermore, we have tested whether in vivo administration of LP-211, a selective 5-HT7R agonist, can rescue learning and behavior in Fmr1 KO mice. In the presence of an adenylate cyclase blocker, mGluR-LTD was slightly enhanced in WT and therefore the difference between mGluR-LTD in WT and Fmr1 KO slices was no longer present. Conversely, activation of adenylate cyclase by either forskolin or Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) completely reversed mGluR-LTD in WT and Fmr1 KO. 5-HT7R activation reversed mGluR-LTD in WT and corrected exaggerated mGluR-LTD in Fmr1 KO; this effect was abolished by blockade of either adenylate cyclase or protein kinase A (PKA). Exposure of hippocampal slices to LP-211 caused an increased phosphorylation of extracellular signal regulated kinase (ERK), an intracellular effector involved in mGluR-LTD, in WT mice. Conversely, this effect was barely detectable in Fmr1 KO mice, suggesting that 5-HT7R-mediated reversal of mGluR-LTD does not require ERK stimulation. Finally, an acute in vivo administration of LP-211 improved novel object recognition (NOR) performance in WT and Fmr1 KO mice and reduced stereotyped behavior in Fmr1 KO mice. Our results indicate that mGluR-LTD in WT and Fmr1 KO slices is bidirectionally modulated in conditions of either reduced or enhanced cAMP formation. Activation of 5-HT7 receptors reverses mGluR-LTD by activation of the cAMP/PKA intracellular pathway. Importantly, a systemic administration of a 5-HT7R agonist to Fmr1 KO mice corrected learning deficits and repetitive behavior. We suggest that selective 5-HT7R agonists might become novel pharmacological tools for FXS therapy.
Keywords: 5-HT7 receptor; PACAP; cyclic AMP; fragile X syndrome; learning; mGluR-LTD; serotonin.
Figures
References
-
- Andreetta F., Carboni L., Grafton G., Jeggo R., Whyment A. D., van den Top M., et al. (2016). Hippocampal 5-HT7 receptors signal phosphorylation of the GluA1 subunit to facilitate AMPA receptor mediated-neurotransmission in vitro and in vivo. Br. J. Pharmacol. 173, 1438–1451. 10.1111/bph.13432 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous