

FACT complex gene duplicates exhibit redundant and non-redundant functions in C. elegans
- PMID: 30336114
- PMCID: PMC6310015
- DOI: 10.1016/j.ydbio.2018.10.002
FACT complex gene duplicates exhibit redundant and non-redundant functions in C. elegans
Abstract
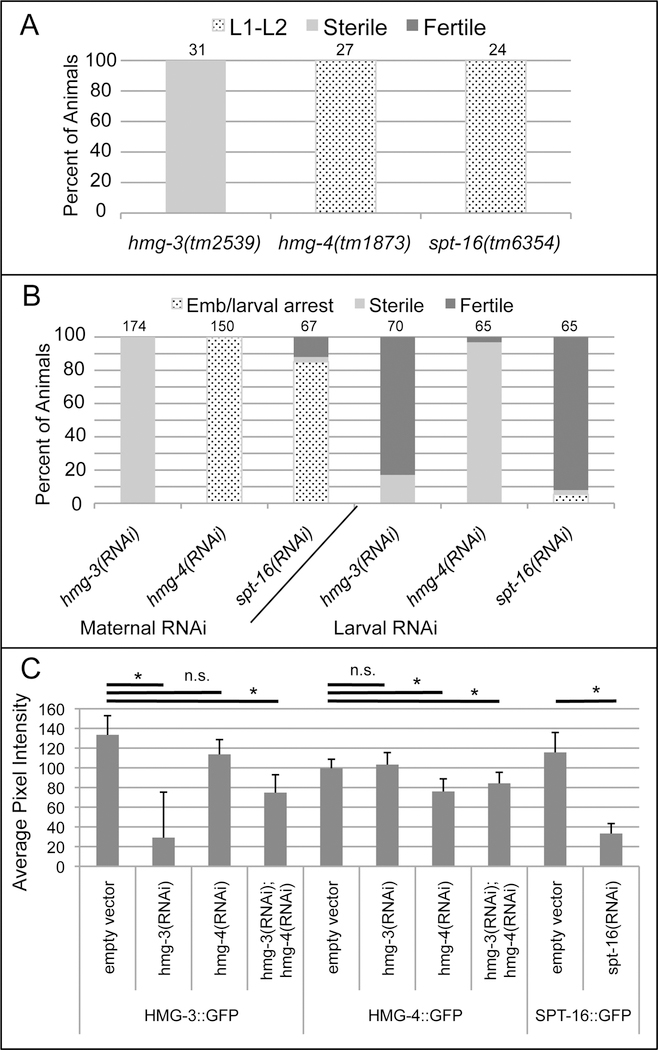
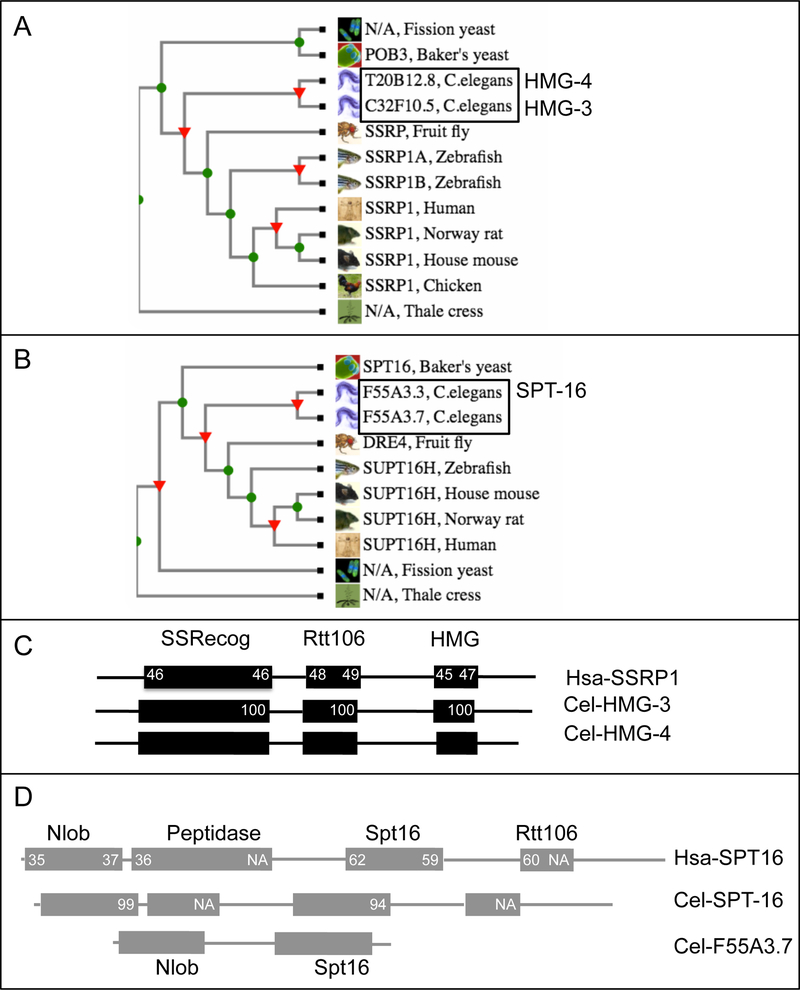
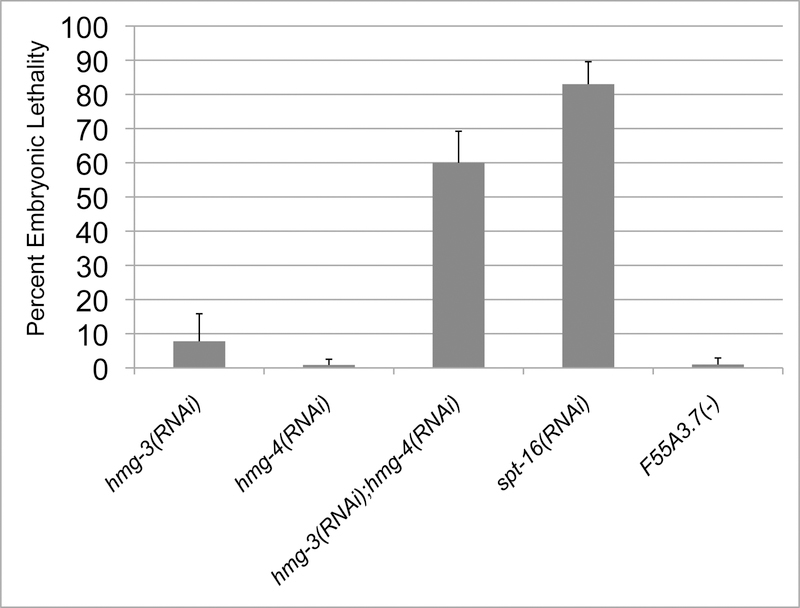
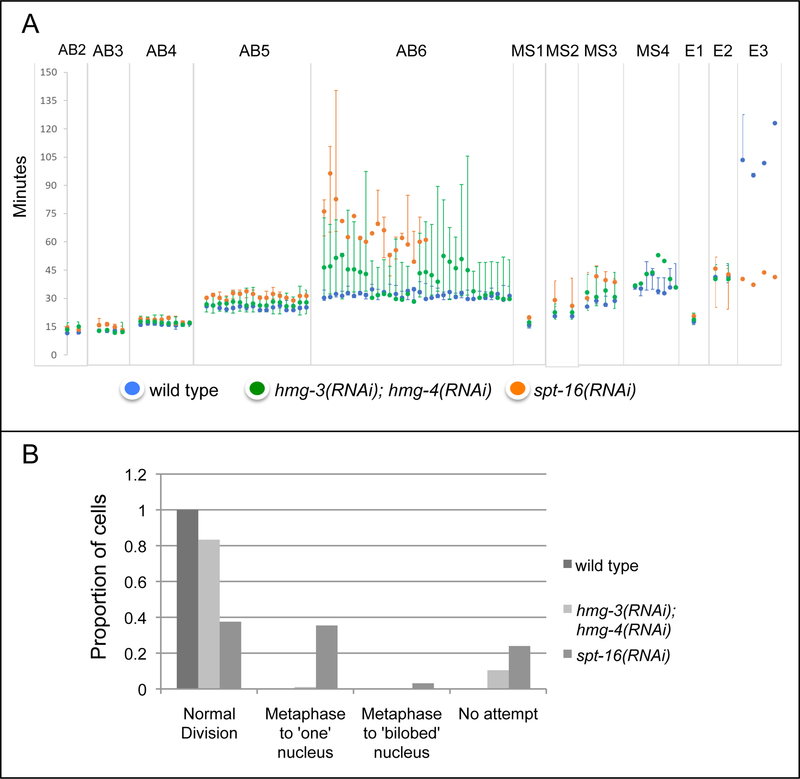
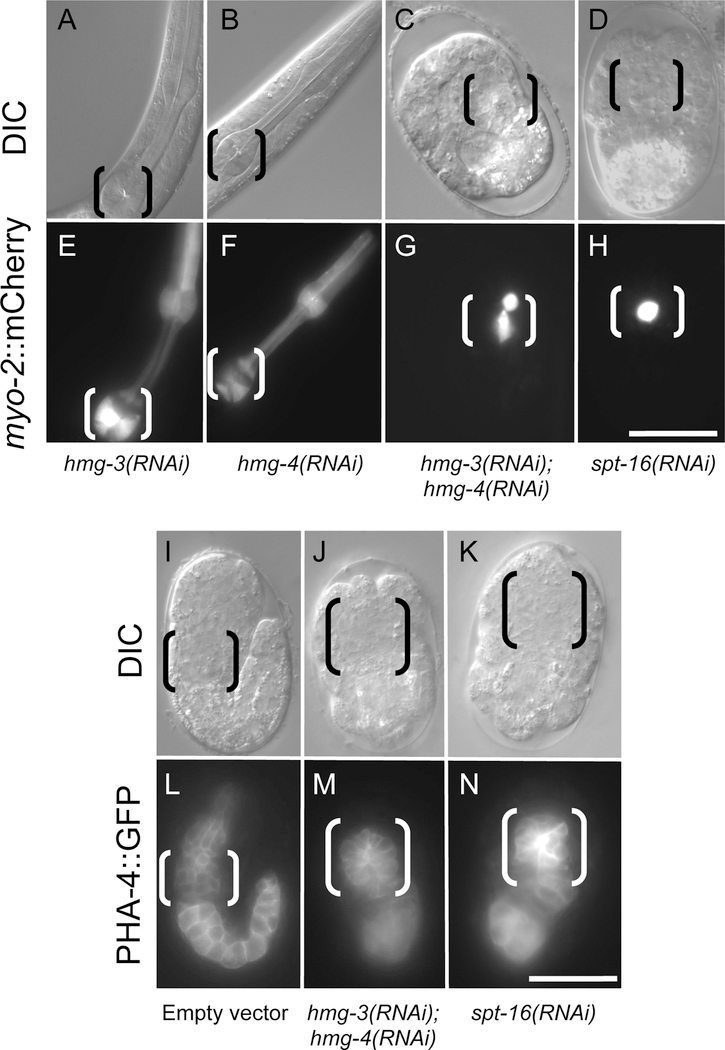
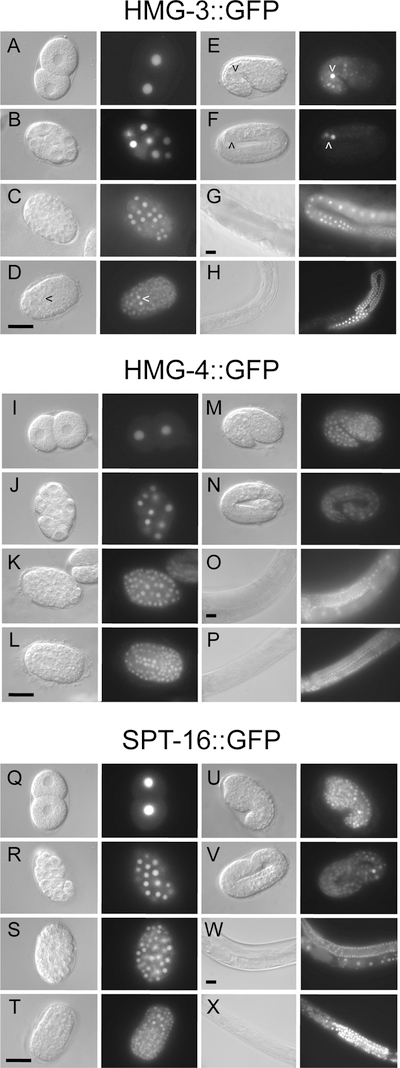
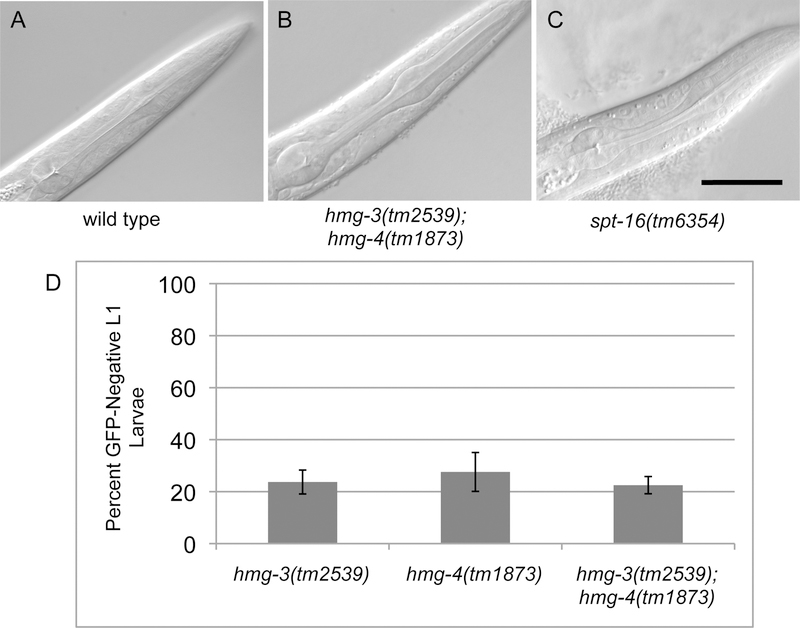
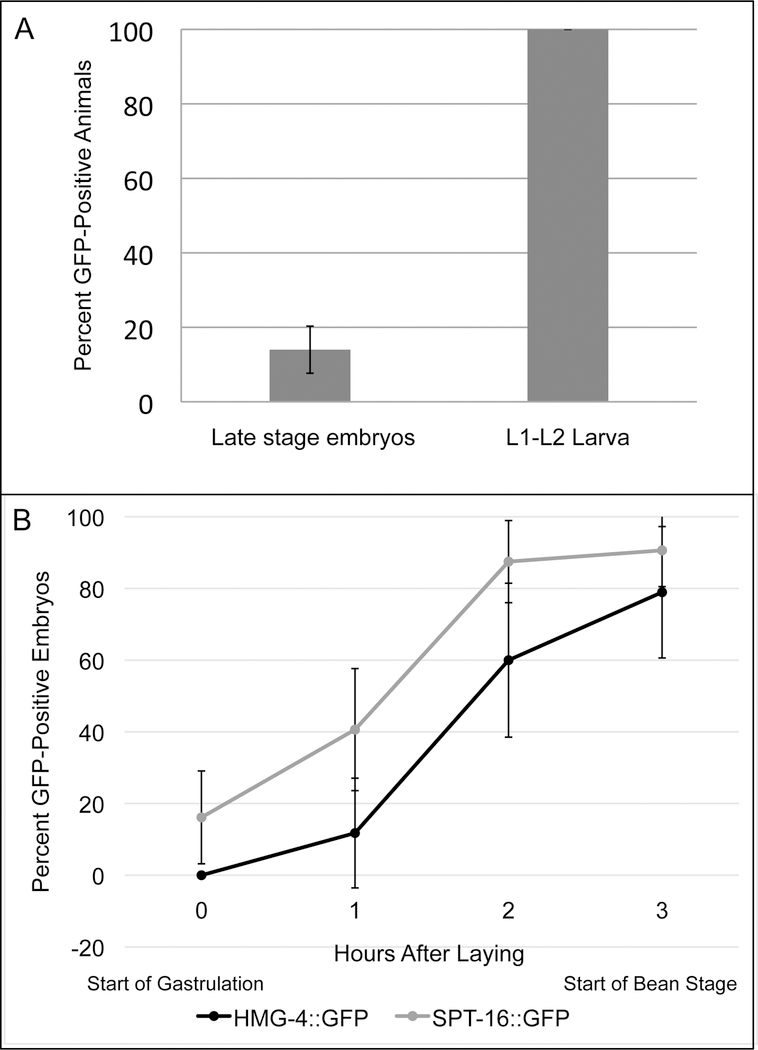
FACT (facilitates chromatin transcription) is a histone chaperone complex important in genomic processes including transcription, DNA replication, and DNA repair. FACT is composed of two proteins, SSRP1 and SPT16, which are highly conserved across eukaryotes. While the mechanisms for FACT in nucleosome reorganization and its relationship to DNA processes is well established, how these roles impact coordination in multicellular animal development are less well understood. Here we characterize the genes encoding FACT complex proteins in the nematode C. elegans. We show that whereas C. elegans includes one SPT16 gene (spt-16), two genes (hmg-3 and hmg-4) encode SSRP1 proteins. Depletion of FACT complex genes interferes with embryonic cell division and cell cycle timing generally, with anterior pharynx development especially sensitive to these defects. hmg-3 and hmg-4 exhibit redundancy for these maternally-provided embryonic functions, but are each uniquely required zygotically for normal germline development. This work provides a framework to study FACT gene function in developmental processes, and identifies that distinct functional requirements for gene duplicates can be manifest within a single tissue.
Keywords: Cell cycle timing; Gene duplication; Pharyngeal development; SPT16; SSRP1.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interest: none
Figures

Similar articles
-
Regulation of chromatin structure and function: insights into the histone chaperone FACT.Cell Cycle. 2021 Mar-Mar;20(5-6):465-479. doi: 10.1080/15384101.2021.1881726. Epub 2021 Feb 16. Cell Cycle. 2021. PMID: 33590780 Free PMC article. Review.
-
Spt16-Pob3 and the HMG protein Nhp6 combine to form the nucleosome-binding factor SPN.EMBO J. 2001 Jul 2;20(13):3506-17. doi: 10.1093/emboj/20.13.3506. EMBO J. 2001. PMID: 11432837 Free PMC article.
-
Functions of FACT in Breaking the Nucleosome and Maintaining Its Integrity at the Single-Nucleosome Level.Mol Cell. 2018 Jul 19;71(2):284-293.e4. doi: 10.1016/j.molcel.2018.06.020. Mol Cell. 2018. PMID: 30029006
-
Histone chaperone FACT coordinates nucleosome interaction through multiple synergistic binding events.J Biol Chem. 2011 Dec 2;286(48):41883-41892. doi: 10.1074/jbc.M111.301465. Epub 2011 Oct 3. J Biol Chem. 2011. PMID: 21969370 Free PMC article.
-
The histone chaperone FACT: structural insights and mechanisms for nucleosome reorganization.J Biol Chem. 2011 May 27;286(21):18369-74. doi: 10.1074/jbc.R110.180778. Epub 2011 Mar 24. J Biol Chem. 2011. PMID: 21454601 Free PMC article. Review.
Cited by
-
Automated profiling of gene function during embryonic development.Cell. 2024 Jun 6;187(12):3141-3160.e23. doi: 10.1016/j.cell.2024.04.012. Epub 2024 May 16. Cell. 2024. PMID: 38759650 Free PMC article.
-
A 4D single-cell protein atlas of transcription factors delineates spatiotemporal patterning during embryogenesis.Nat Methods. 2021 Aug;18(8):893-902. doi: 10.1038/s41592-021-01216-1. Epub 2021 Jul 26. Nat Methods. 2021. PMID: 34312566
-
Gene expression profiling of epidermal cell types in C. elegans using Targeted DamID.Development. 2021 Sep 1;148(17):dev199452. doi: 10.1242/dev.199452. Epub 2021 Sep 3. Development. 2021. PMID: 34397094 Free PMC article.
-
Regulation of chromatin structure and function: insights into the histone chaperone FACT.Cell Cycle. 2021 Mar-Mar;20(5-6):465-479. doi: 10.1080/15384101.2021.1881726. Epub 2021 Feb 16. Cell Cycle. 2021. PMID: 33590780 Free PMC article. Review.
-
Serotonin signaling by maternal neurons upon stress ensures progeny survival.Elife. 2020 Apr 23;9:e55246. doi: 10.7554/eLife.55246. Elife. 2020. PMID: 32324136 Free PMC article.
References
-
- Abe T, Sugimura K, Hosono Y, Takami Y, Akita M, Yoshimura A, Tada S, Nakayama T, Murofushi H, Okumura K, Takeda S, Horikoshi M, Seki M, Enomoto T, 2011. The histone chaperone facilitates chromatin transcription (FACT) protein maintains normal replication fork rates. J. Biol. Chem 286, 30504–30512. 10.1074/jbc.M111.264721 - DOI - PMC - PubMed
-
- Baugh LR, Hill AA, Slonim DK, Brown EL, Hunter CP, 2003. Composition and dynamics of the Caenorhabditis elegans early embryonic transcriptome. Dev. Camb. Engl 130, 889–900. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
