

Parts-Prospecting for a High-Efficiency Thiamin Thiazole Biosynthesis Pathway
- PMID: 30337452
- PMCID: PMC6393793
- DOI: 10.1104/pp.18.01085
Parts-Prospecting for a High-Efficiency Thiamin Thiazole Biosynthesis Pathway
Abstract
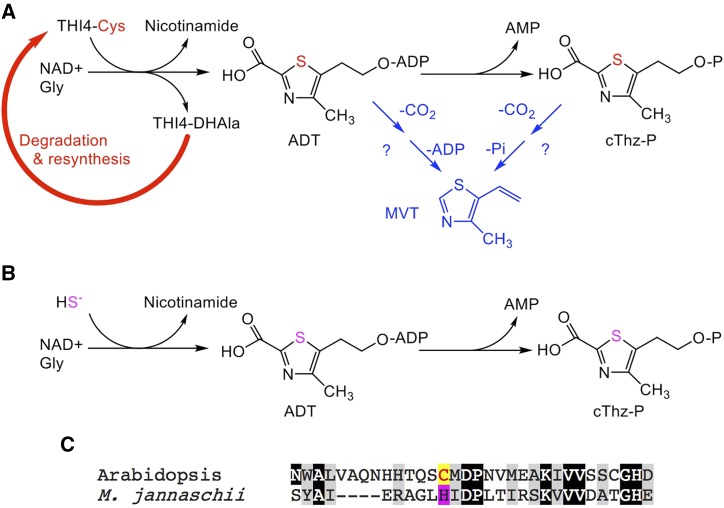
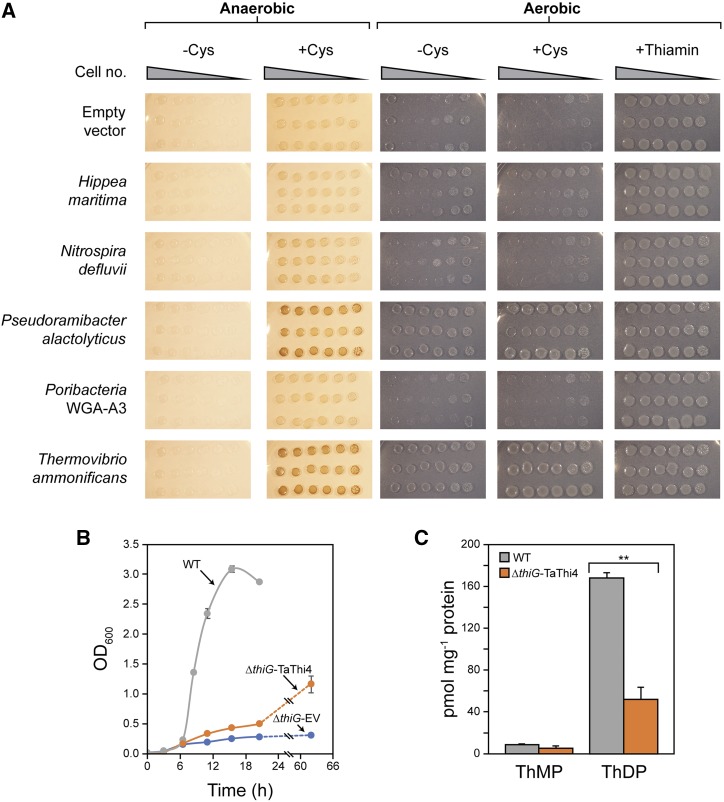
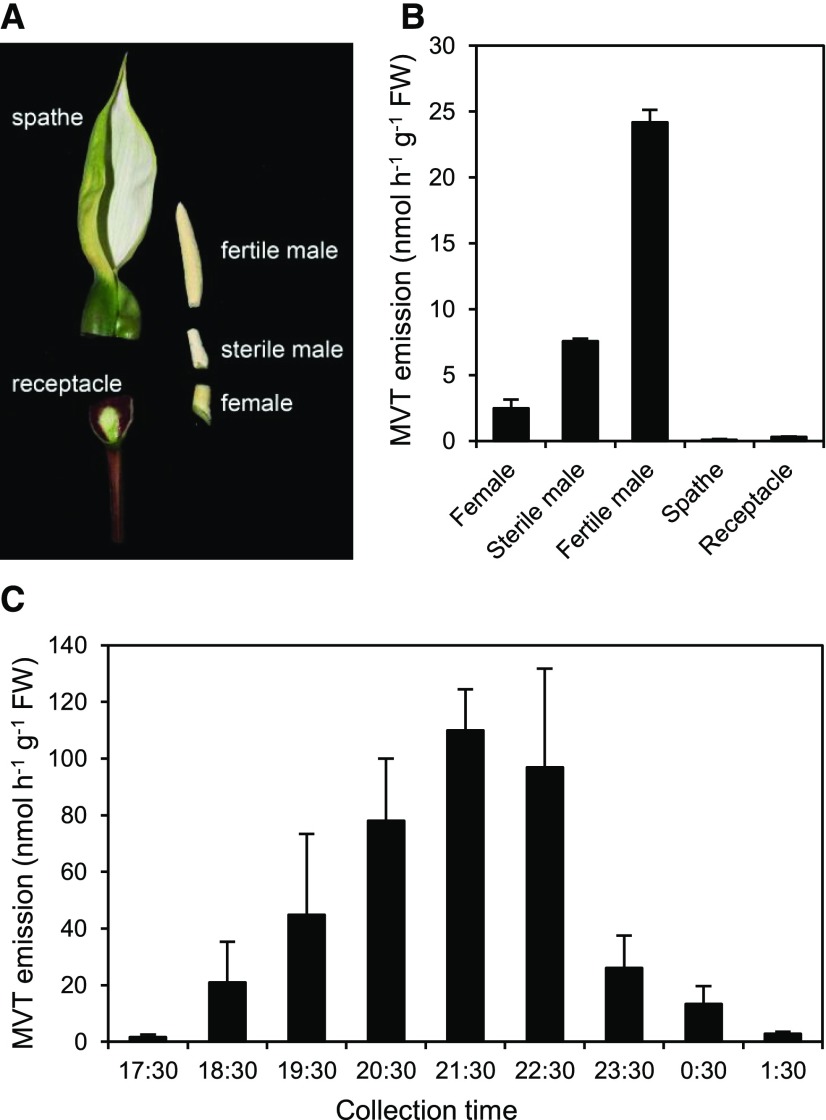
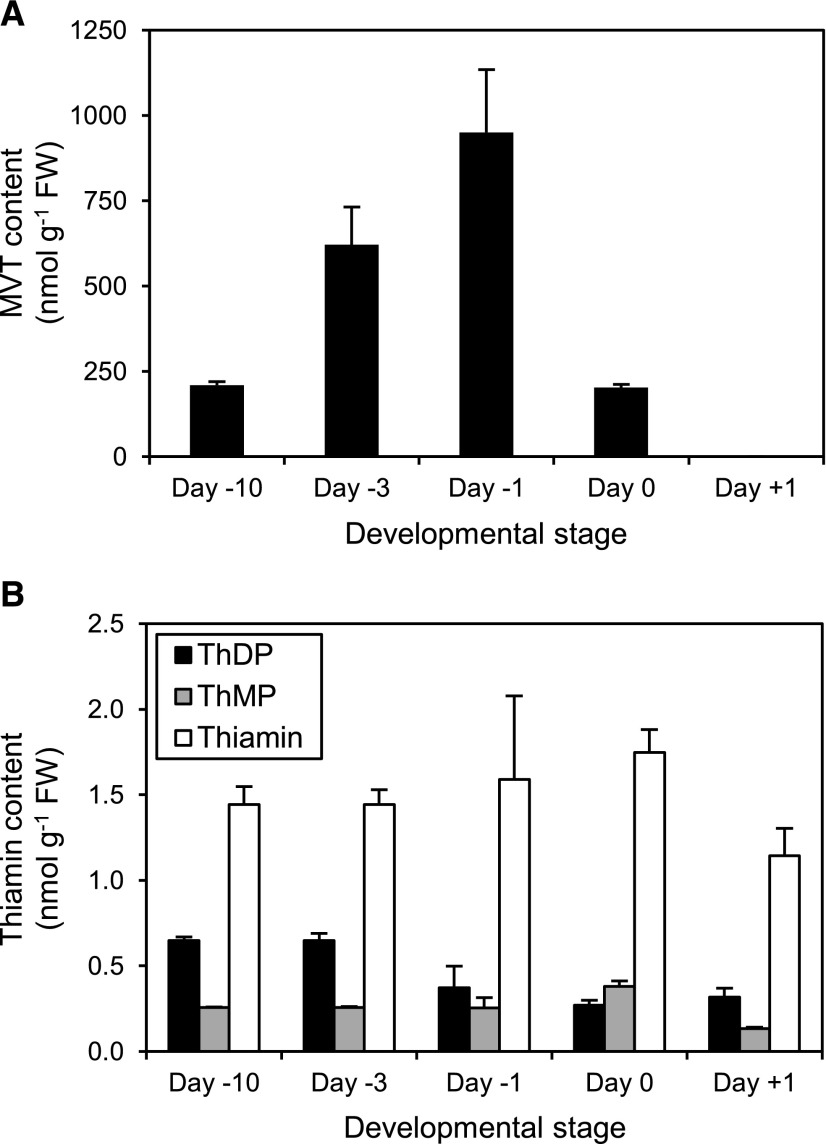
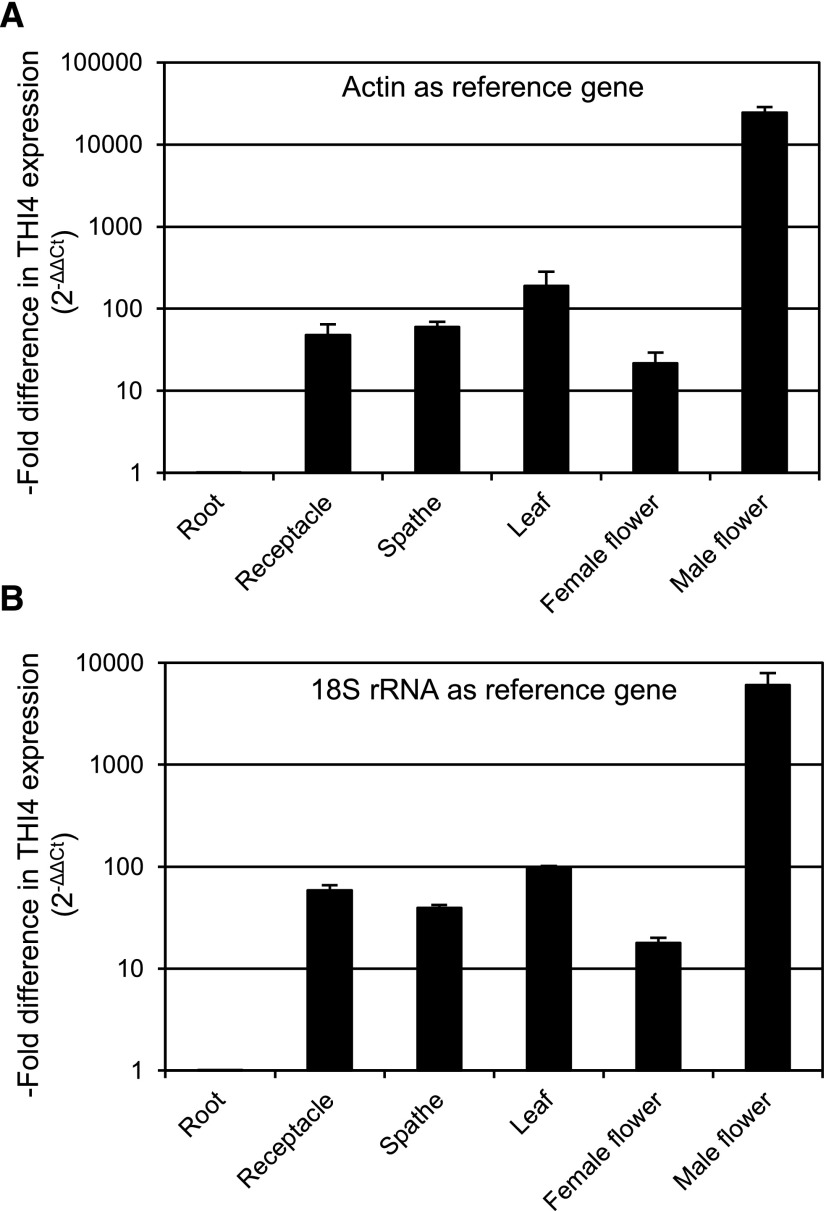
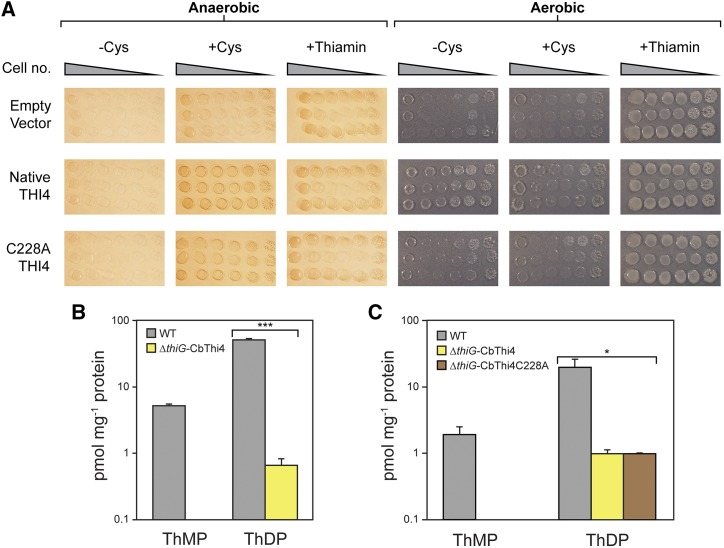
Plants synthesize the thiazole precursor of thiamin (cThz-P) via THIAMIN4 (THI4), a suicide enzyme that mediates one reaction cycle and must then be degraded and resynthesized. It has been estimated that this THI4 turnover consumes 2% to 12% of the maintenance energy budget and that installing an energy-efficient alternative pathway could substantially increase crop yield potential. Available data point to two natural alternatives to the suicidal THI4 pathway: (i) nonsuicidal prokaryotic THI4s that lack the active-site Cys residue on which suicide activity depends, and (ii) an uncharacterized thiazole synthesis pathway in flowers of the tropical arum lily Caladium bicolor that enables production and emission of large amounts of the cThz-P analog 4-methyl-5-vinylthiazole (MVT). We used functional complementation of an Escherichia coli ΔthiG strain to identify a nonsuicidal bacterial THI4 (from Thermovibrio ammonificans) that can function in conditions like those in plant cells. We explored whether C. bicolor synthesizes MVT de novo via a novel route, via a suicidal or a nonsuicidal THI4, or by catabolizing thiamin. Analysis of developmental changes in MVT emission, extractable MVT, thiamin level, and THI4 expression indicated that C. bicolor flowers make MVT de novo via a massively expressed THI4 and that thiamin is not involved. Functional complementation tests indicated that C. bicolor THI4, which has the active-site Cys needed to operate suicidally, may be capable of suicidal and - in hypoxic conditions - nonsuicidal operation. T. ammonificans and C. bicolor THI4s are thus candidate parts for rational redesign or directed evolution of efficient, nonsuicidal THI4s for use in crop improvement.
© 2019 American Society of Plant Biologists. All rights reserved.
Figures
References
-
- Ap Rees T, Wright BW, Fuller WA (1977) Measurements of starch breakdown as estimates of glycolysis during thermogenesis by the spadix of Arum maculatum L. Planta 134: 53–56 - PubMed
-
- Belanger FC, Leustek T, Chu B, Kriz AL (1995) Evidence for the thiamine biosynthetic pathway in higher-plant plastids and its developmental regulation. Plant Mol Biol 29: 809–821 - PubMed
-
- Bessman MJ, Frick DN, O’Handley SF (1996) The MutT proteins or “Nudix” hydrolases, a family of versatile, widely distributed, “housecleaning” enzymes. J Biol Chem 271: 25059–25062 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
