

The enigmatic ATP supply of the endoplasmic reticulum
- PMID: 30338910
- PMCID: PMC6446729
- DOI: 10.1111/brv.12469
The enigmatic ATP supply of the endoplasmic reticulum
Abstract
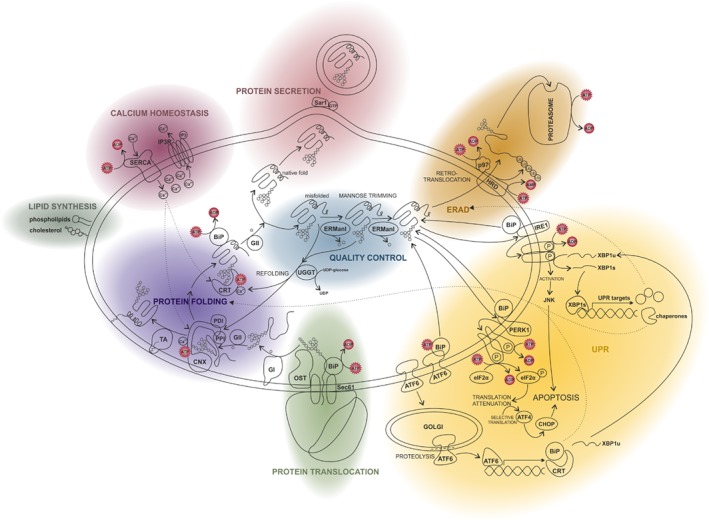
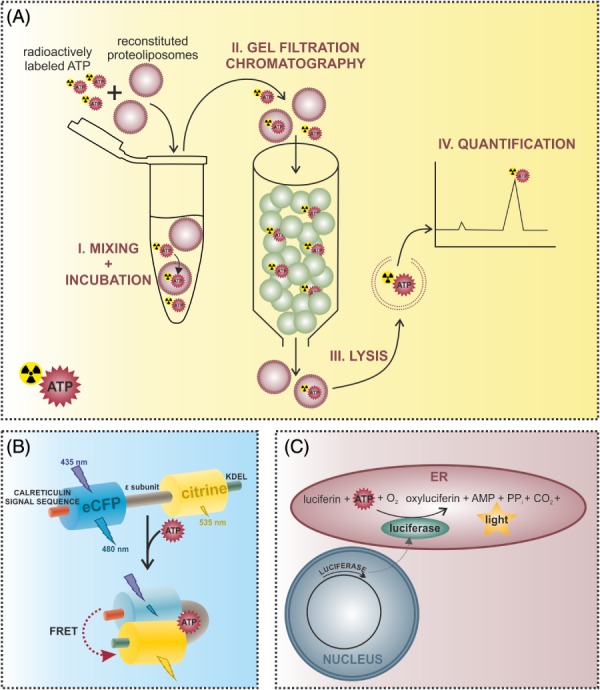
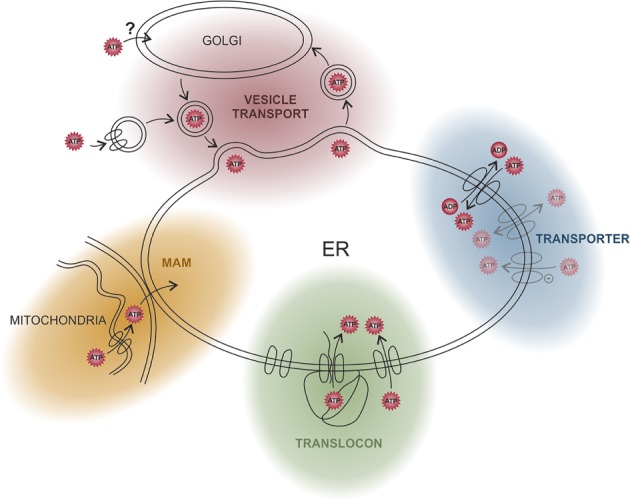
The endoplasmic reticulum (ER) is a functionally and morphologically complex cellular organelle largely responsible for a variety of crucial functions, including protein folding, maturation and degradation. Furthermore, the ER plays an essential role in lipid biosynthesis, dynamic Ca2+ storage, and detoxification. Malfunctions in ER-related processes are responsible for the genesis and progression of many diseases, such as heart failure, cancer, neurodegeneration and metabolic disorders. To fulfill many of its vital functions, the ER relies on a sufficient energy supply in the form of adenosine-5'-triphosphate (ATP), the main cellular energy source. Despite landmark discoveries and clarification of the functional principles of ER-resident proteins and key ER-related processes, the mechanism underlying ER ATP transport remains somewhat enigmatic. Here we summarize ER-related ATP-consuming processes and outline our knowledge about the nature and function of the ER energy supply.
Keywords: ATP; ATP transporter; ER stress; ERAD; endoplasmic reticulum (ER); protein quality control; secretory pathway; unfolded protein response (UPR).
© 2018 The Authors. Biological Reviews published by John Wiley & Sons Ltd on behalf of Cambridge Philosophical Society.
Figures
References
-
- Abele, R. & Tampé, R. (2011). The TAP translocation machinery in adaptive immunity and viral escape mechanisms. Essays in Biochemistry 50, 249–264. - PubMed
-
- Ainscow, E. K. & Rutter, G. A. (2002). Glucose‐stimulated oscillations in free cytosolic ATP concentration imaged in single islet β‐cells evidence for a Ca2+‐dependent mechanism. Diabetes 51, S162–S170. - PubMed
-
- Allan, B. B. , Moyer, B. D. & Balch, W. E. (2000). Rab1 recruitment of p115 into a cis‐SNARE complex: programming budding COPII vesicles for fusion. Science (New York, N.Y.) 289, 444–448. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
