

Harnessing accurate non-homologous end joining for efficient precise deletion in CRISPR/Cas9-mediated genome editing
- PMID: 30340517
- PMCID: PMC6195759
- DOI: 10.1186/s13059-018-1518-x
Harnessing accurate non-homologous end joining for efficient precise deletion in CRISPR/Cas9-mediated genome editing
Abstract
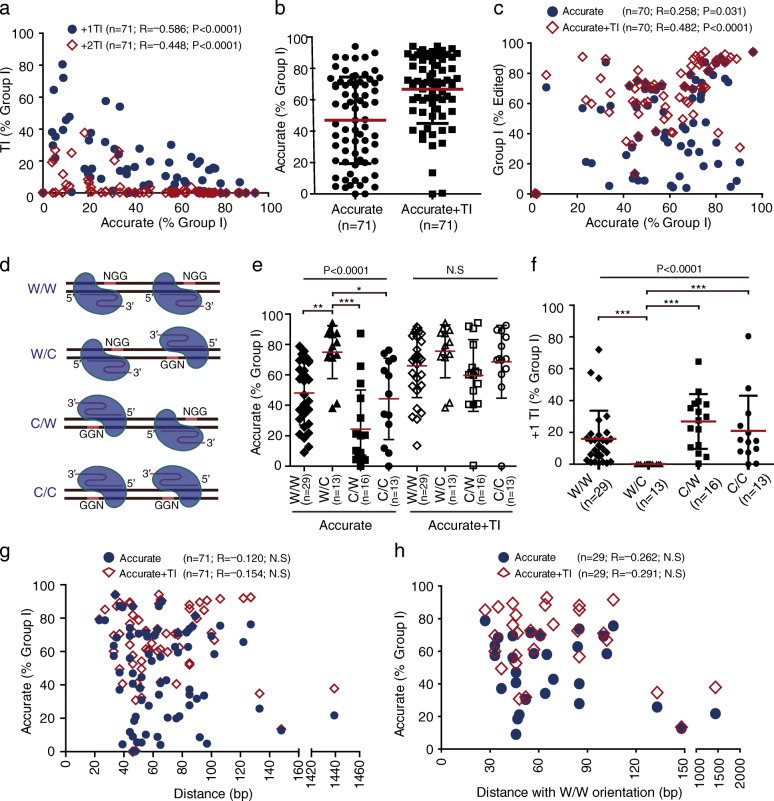
Background: Many applications of CRISPR/Cas9-mediated genome editing require Cas9-induced non-homologous end joining (NHEJ), which was thought to be error prone. However, with directly ligatable ends, Cas9-induced DNA double strand breaks may be repaired preferentially by accurate NHEJ.
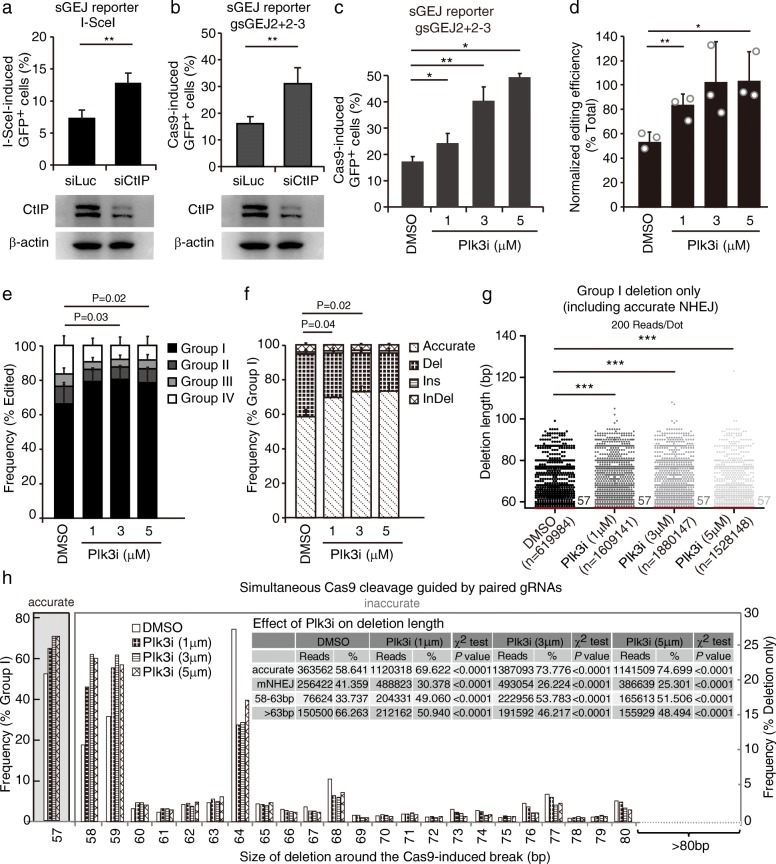
Results: In the repair of two adjacent double strand breaks induced by paired Cas9-gRNAs at 71 genome sites, accurate NHEJ accounts for about 50% of NHEJ events. This paired Cas9-gRNA approach underestimates the level of accurate NHEJ due to frequent + 1 templated insertions, which can be avoided by the predefined Watson/Crick orientation of protospacer adjacent motifs (PAMs). The paired Cas9-gRNA strategy also provides a flexible, reporter-less approach for analyzing both accurate and mutagenic NHEJ in cells and in vivo, and it has been validated in cells deficient for XRCC4 and in mouse liver. Due to high frequencies of precise deletions of defined "3n"-, "3n + 1"-, or "3n + 2"-bp length, accurate NHEJ is used to improve the efficiency and homogeneity of gene knockouts and targeted in-frame deletions. Compared to "3n + 1"-bp, "3n + 2"-bp can overcome + 1 templated insertions to increase the frequency of out-of-frame mutations. By applying paired Cas9-gRNAs to edit MDC1 and key 53BP1 domains, we are able to generate predicted, precise deletions for functional analysis. Lastly, a Plk3 inhibitor promotes NHEJ with bias towards accurate NHEJ, providing a chemical approach to improve genome editing requiring precise deletions.
Conclusions: NHEJ is inherently accurate in repair of Cas9-induced DNA double strand breaks and can be harnessed to improve CRISPR/Cas9 genome editing requiring precise deletion of a defined length.
Keywords: Accurate NHEJ; CRISPR/Cas9; Genome editing; Knockout; Paired gRNAs; Precise deletion; Targeted in-frame deletion; Templated insertions.
Conflict of interest statement
Ethics approval and consent to participate
All animal procedures were reviewed and revised by the Animal Welfare and Ethical Review Body at Zhejiang University, with approval number ZJU2015–378-01.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
