

Mutations in Peptidoglycan Synthesis Gene ponA Improve Electrotransformation Efficiency of Corynebacterium glutamicum ATCC 13869
- PMID: 30341076
- PMCID: PMC6275343
- DOI: 10.1128/AEM.02225-18
Mutations in Peptidoglycan Synthesis Gene ponA Improve Electrotransformation Efficiency of Corynebacterium glutamicum ATCC 13869
Abstract
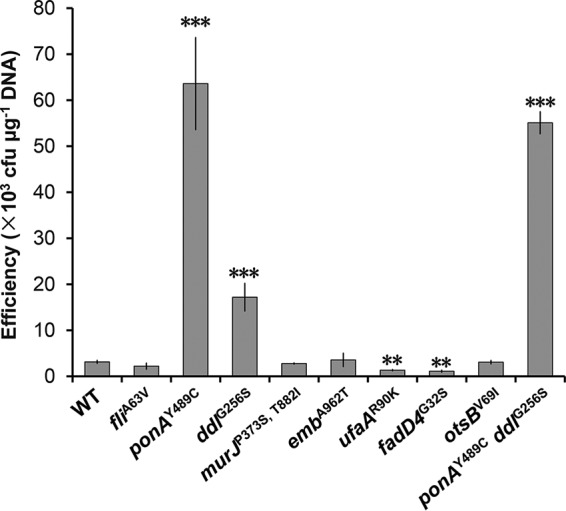
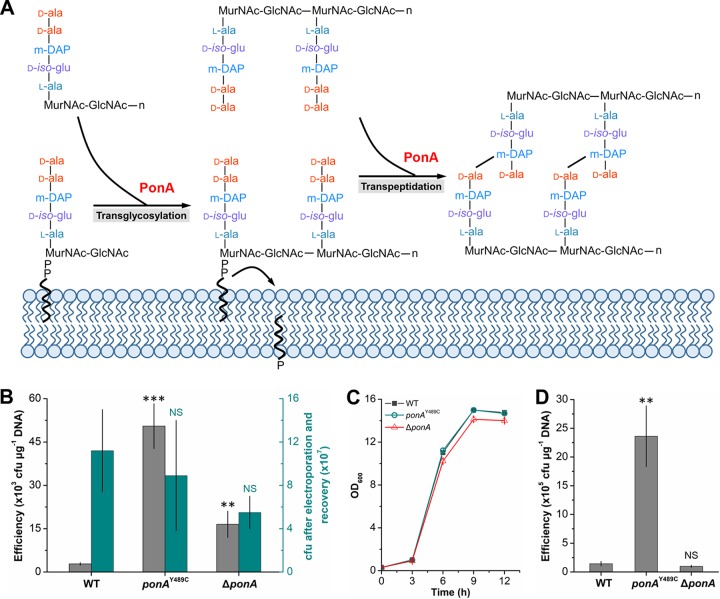
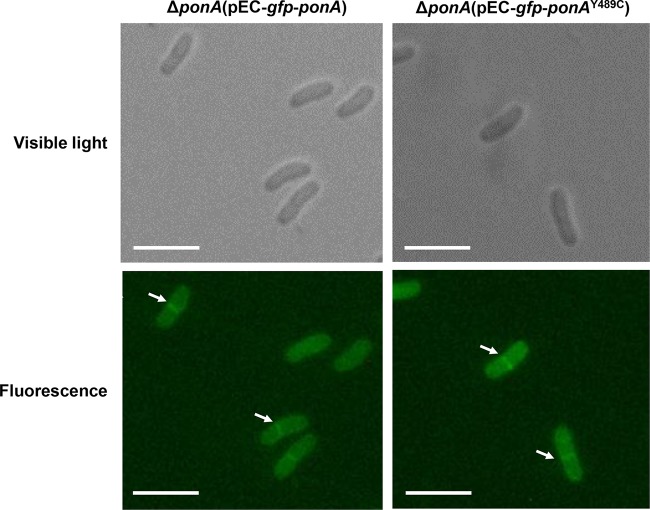
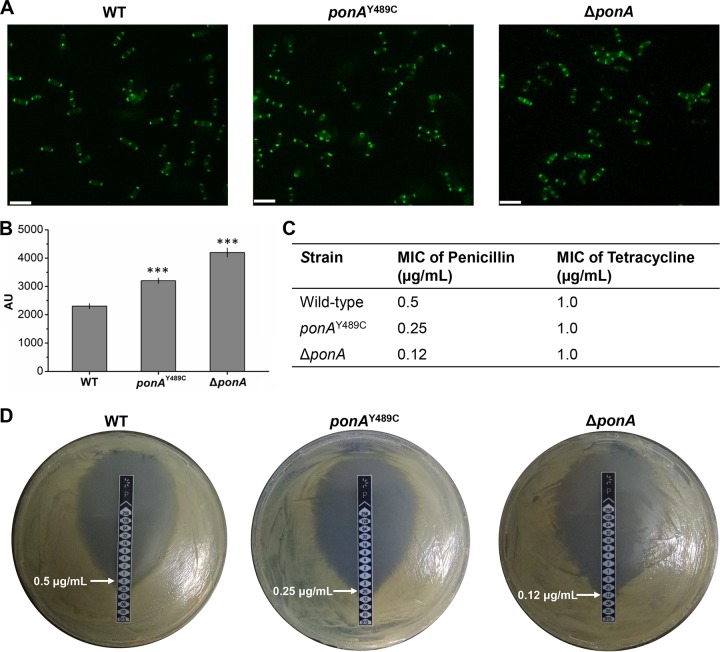
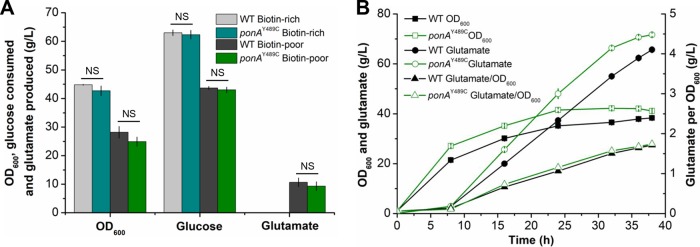
Corynebacterium glutamicum is frequently engineered to serve as a versatile platform and model microorganism. However, due to its complex cell wall structure, transformation of C. glutamicum with exogenous DNA is inefficient. Although efforts have been devoted to improve the transformation efficiency by using cell wall-weakening agents, direct genetic engineering of cell wall synthesis for enhancing cell competency has not been explored thus far. Herein, we reported that engineering of peptidoglycan synthesis could significantly increase the transformation efficiency of C. glutamicum Comparative analysis of C. glutamicum wild-type strain ATCC 13869 and a mutant with high electrotransformation efficiency revealed nine mutations in eight cell wall synthesis-related genes. Among them, the Y489C mutation in bifunctional peptidoglycan glycosyltransferase/peptidoglycan dd-transpeptidase PonA dramatically increased the electrotransformation of strain ATCC 13869 by 19.25-fold in the absence of cell wall-weakening agents, with no inhibition on growth. The Y489C mutation had no effect on the membrane localization of PonA but affected the peptidoglycan structure. Deletion of the ponA gene led to more dramatic changes to the peptidoglycan structure but only increased the electrotransformation by 4.89-fold, suggesting that appropriate inhibition of cell wall synthesis benefited electrotransformation more. Finally, we demonstrated that the PonAY489C mutation did not cause constitutive or enhanced glutamate excretion, making its permanent existence in C. glutamicum ATCC 13869 acceptable. This study demonstrates that genetic engineering of genes involved in cell wall synthesis, especially peptidoglycan synthesis, is a promising strategy to improve the electrotransformation efficiency of C. glutamicumIMPORTANCE Metabolic engineering and synthetic biology are now the key enabling technologies for manipulating microorganisms to suit the practical outcomes desired by humankind. The introduction of exogenous DNA into cells is an indispensable step for this purpose. However, some microorganisms, including the important industrial workhorse Corynebacterium glutamicum, possess a complex cell wall structure to shield cells against exogenous DNA. Although genes responsible for cell wall synthesis in C. glutamicum are known, engineering of related genes to improve cell competency has not been explored yet. In this study, we demonstrate that mutations in cell wall synthesis genes can significantly improve the electrotransformation efficiency of C. glutamicum Notably, the Y489C mutation in bifunctional peptidoglycan glycosyltransferase/peptidoglycan dd-transpeptidase PonA increased electrotransformation efficiency by 19.25-fold by affecting peptidoglycan synthesis.
Keywords: Corynebacterium glutamicum; electrotransformation; peptidoglycan; ponA.
Copyright © 2018 American Society for Microbiology.
Figures
References
-
- Kalinowski J, Bathe B, Bartels D, Bischoff N, Bott M, Burkovski A, Dusch N, Eggeling L, Eikmanns BJ, Gaigalat L, Goesmann A, Hartmann M, Huthmacher K, Kramer R, Linke B, McHardy AC, Meyer F, Mockel B, Pfefferle W, Puhler A, Rey DA, Ruckert C, Rupp O, Sahm H, Wendisch VF, Wiegrabe I, Tauch A. 2003. The complete Corynebacterium glutamicum ATCC 13032 genome sequence and its impact on the production of l-aspartate-derived amino acids and vitamins. J Biotechnol 104:5–25. doi:10.1016/S0168-1656(03)00154-8. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
