

Aberrant PRDM9 expression impacts the pan-cancer genomic landscape
- PMID: 30341163
- PMCID: PMC6211651
- DOI: 10.1101/gr.231696.117
Aberrant PRDM9 expression impacts the pan-cancer genomic landscape
Abstract
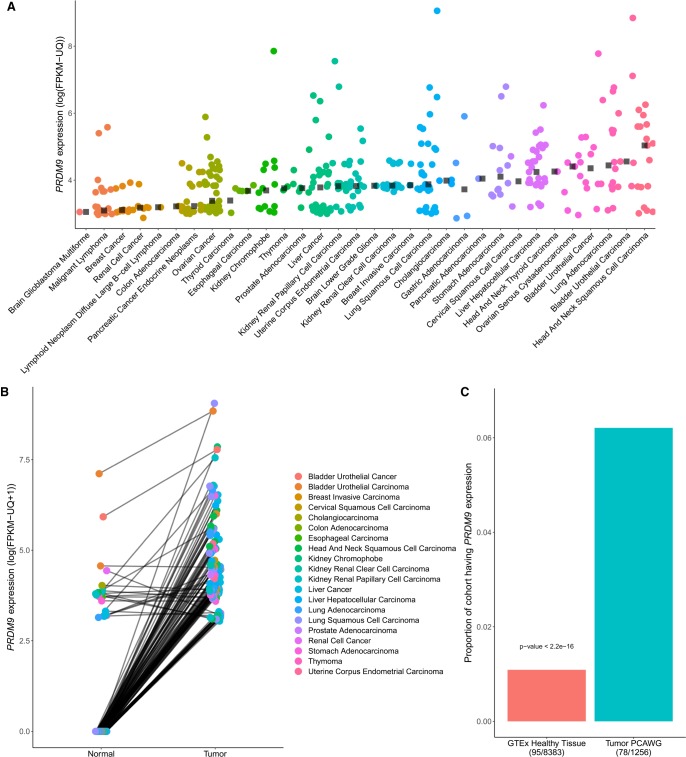
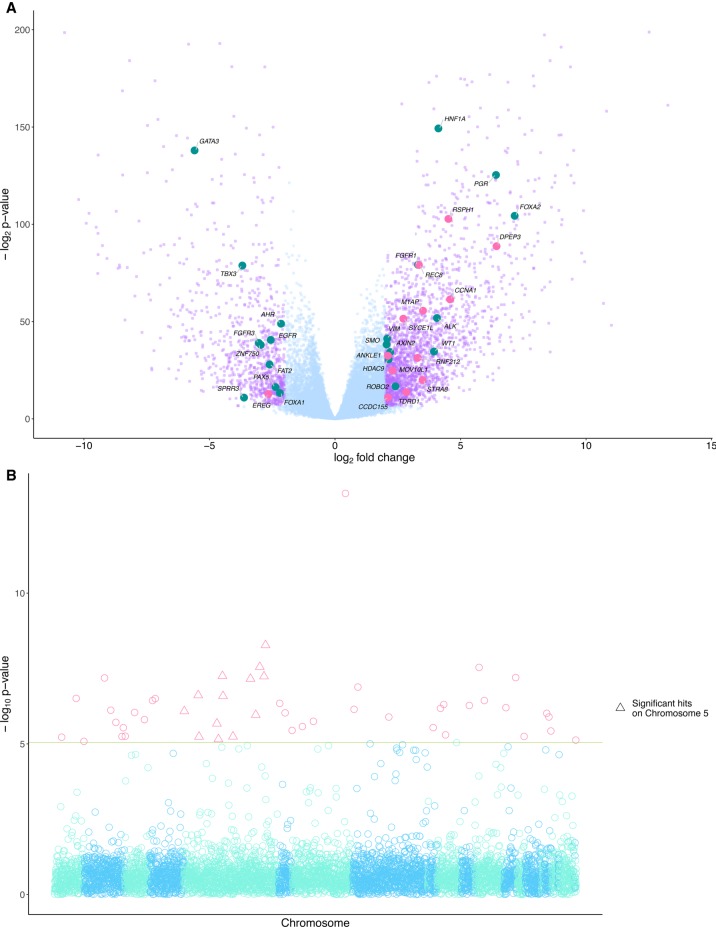
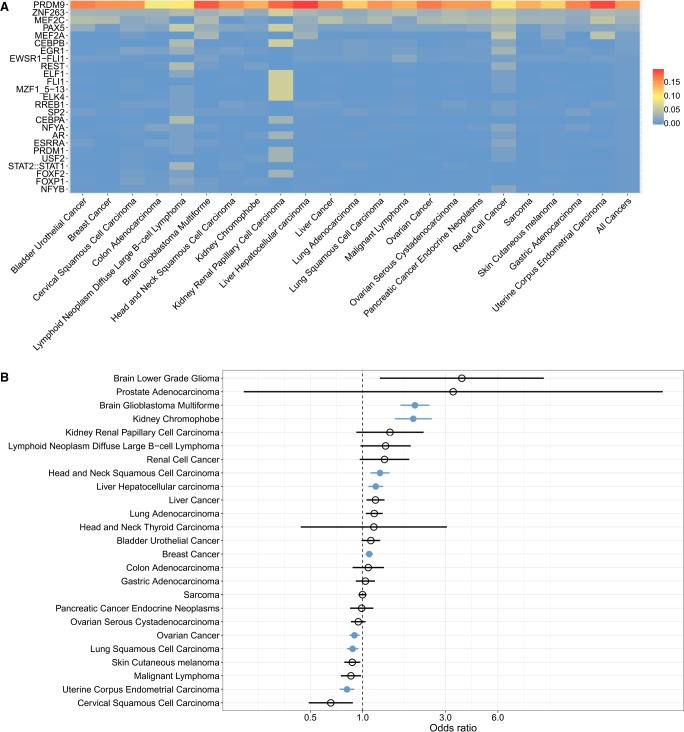
The binding of PRDM9 to chromatin is a key step in the induction of DNA double-strand breaks associated with meiotic recombination hotspots; it is normally expressed solely in germ cells. We interrogated 1879 cancer samples in 39 different cancer types and found that PRDM9 is unexpectedly expressed in 20% of these tumors even after stringent gene homology correction. The expression levels of PRDM9 in tumors are significantly higher than those found in healthy neighboring tissues and in healthy nongerm tissue databases. Recurrently mutated regions located within 5 Mb of the PRDM9 loci, as well as differentially expressed genes in meiotic pathways, correlate with PRDM9 expression. In samples with aberrant PRDM9 expression, structural variant breakpoints frequently neighbor the DNA motif recognized by PRDM9, and there is an enrichment of structural variants at sites of known meiotic PRDM9 activity. This study is the first to provide evidence of an association between aberrant expression of the meiosis-specific gene PRDM9 with genomic instability in cancer.
© 2018 Ang Houle et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous