

DHX9 helicase promotes R-loop formation in cells with impaired RNA splicing
- PMID: 30341290
- PMCID: PMC6195550
- DOI: 10.1038/s41467-018-06677-1
DHX9 helicase promotes R-loop formation in cells with impaired RNA splicing
Abstract
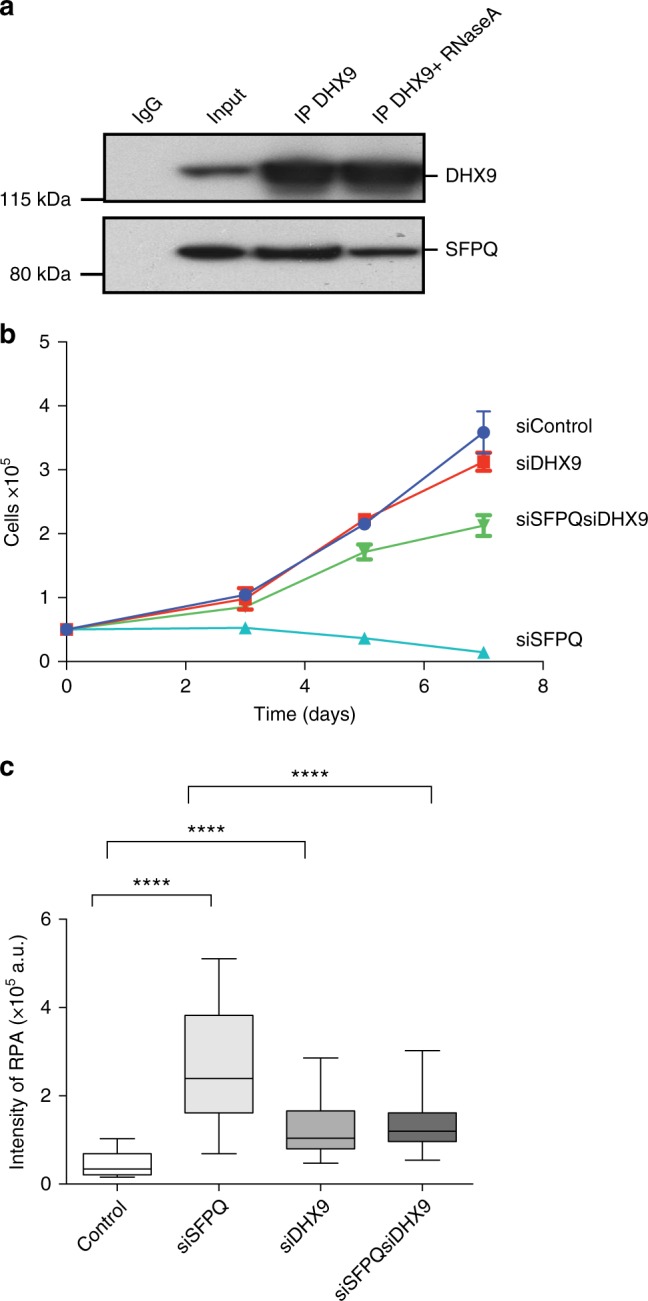
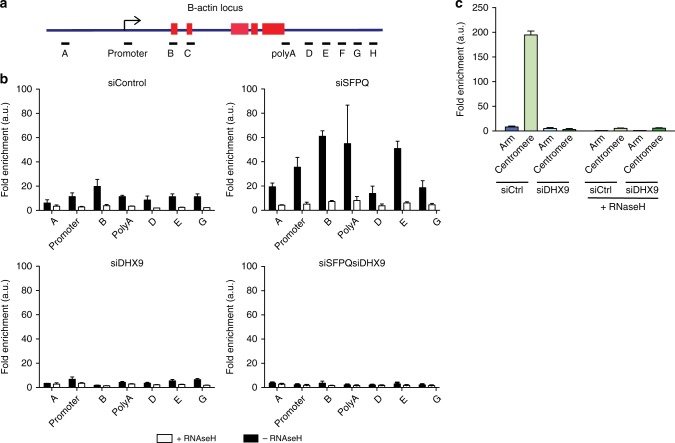
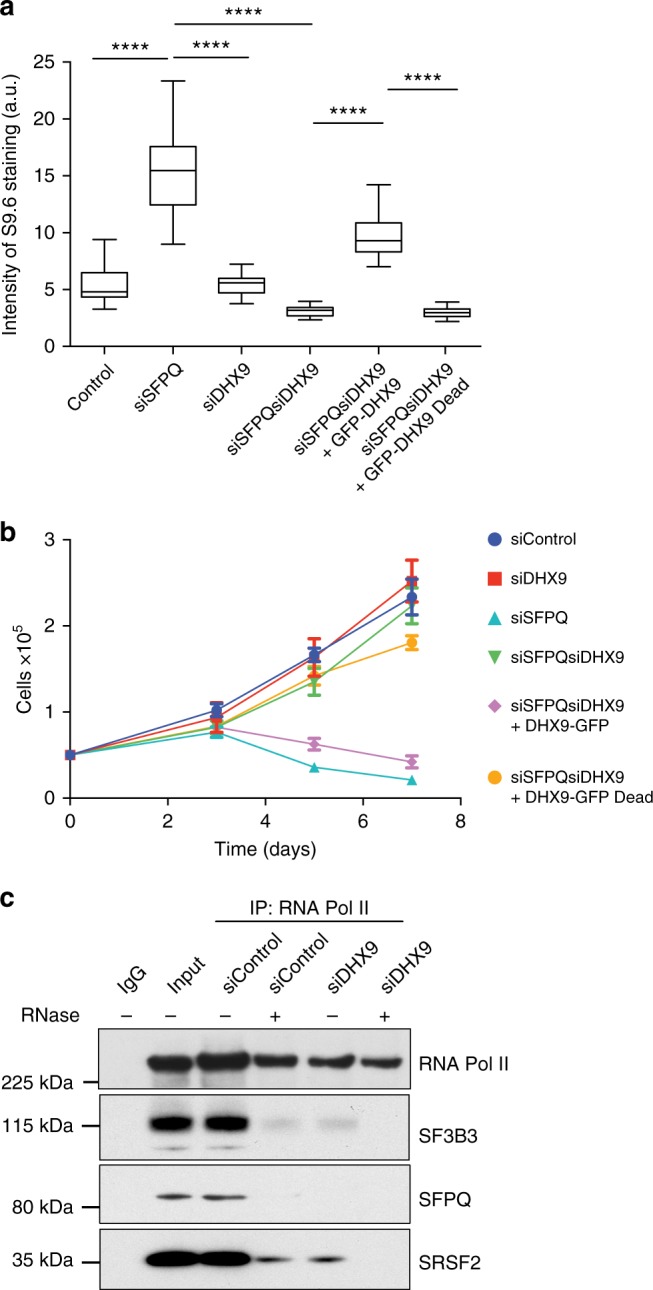
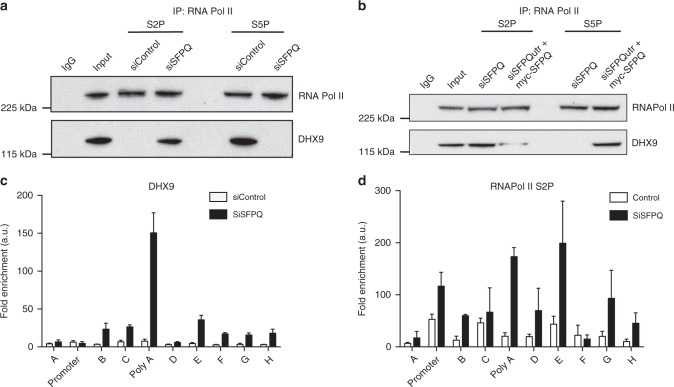
R-loops are stable nucleic acid structures that have important physiological functions, but which also pose a significant threat to genomic stability. Increased R-loops cause replication stress and chromosome fragility and have been associated with diseases such as neurodegeneration and cancer. Although excessive R-loops are a feature of cells that are defective in RNA processing, what causes them to form is unclear. Here, we demonstrate that DHX9 (RNA helicase A) promotes the formation of pathological and non-pathological R-loops. In the absence of splicing factors, formation of R-loops correlates with the prolonged association of DHX9 with RNA Polymerase II (RNA Pol II). This leads to the production of DNA-RNA hybrid, which traps RNA Pol II on chromatin with the potential to block DNA replication. Our data provide a molecular mechanism for the formation of R-loops that is relevant to neurodegenerative diseases and cancers in which deregulated RNA processing is a feature.
Conflict of interest statement
The authors declare no competing interests.
Figures
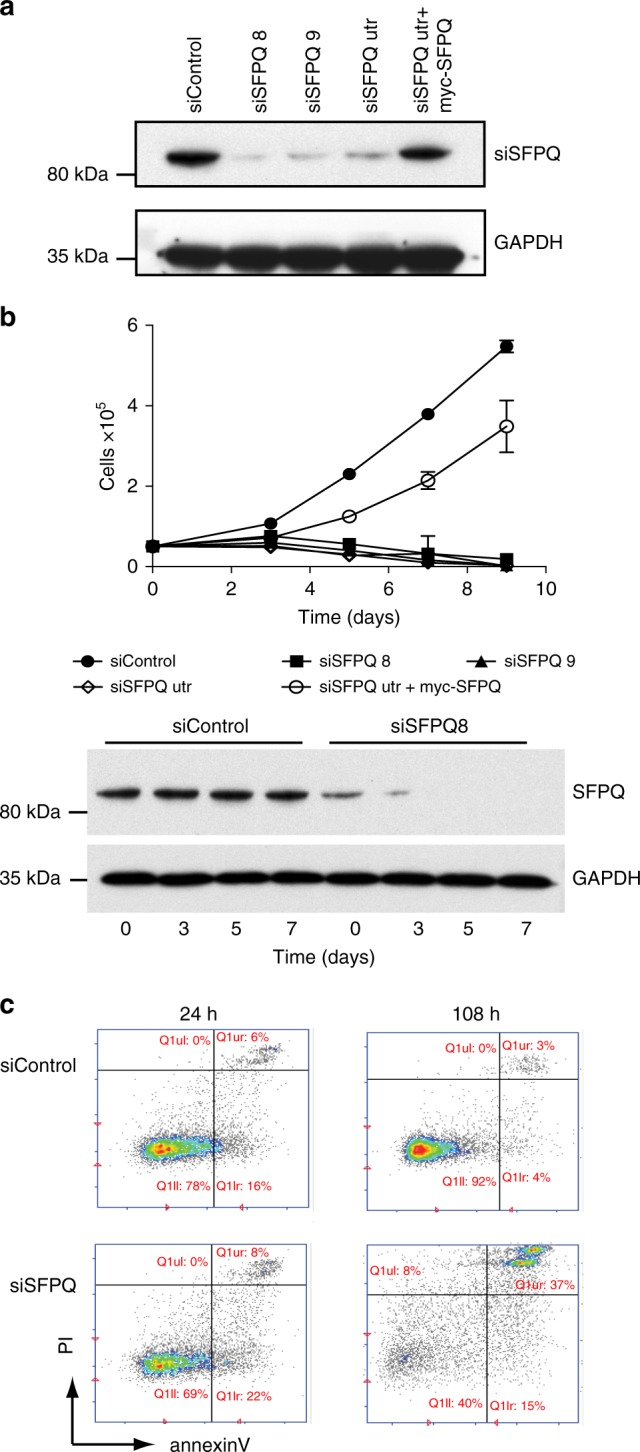
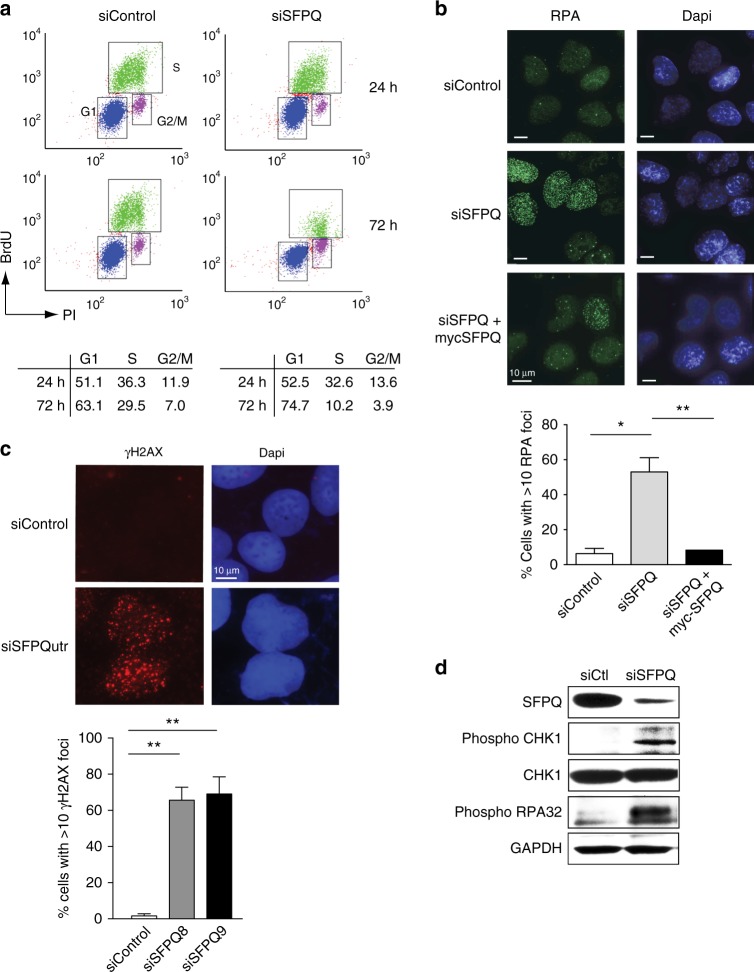
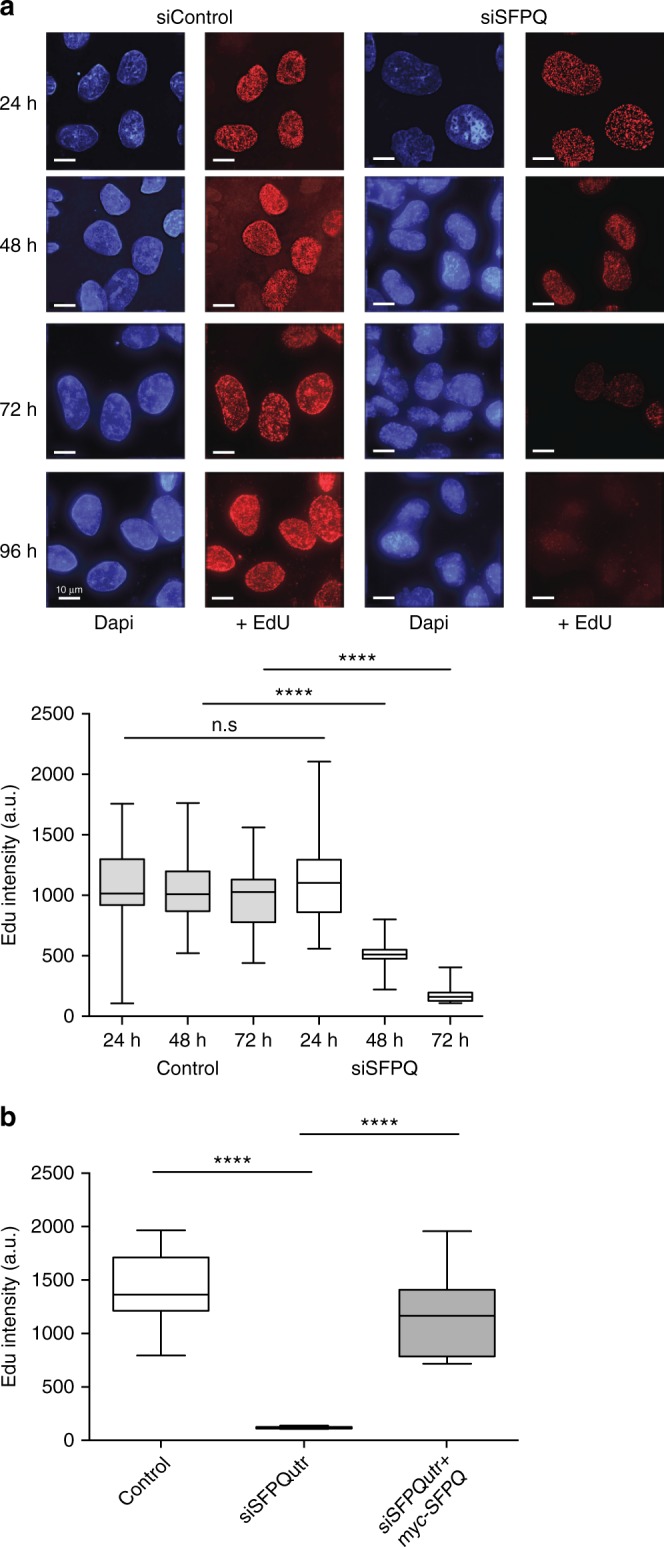
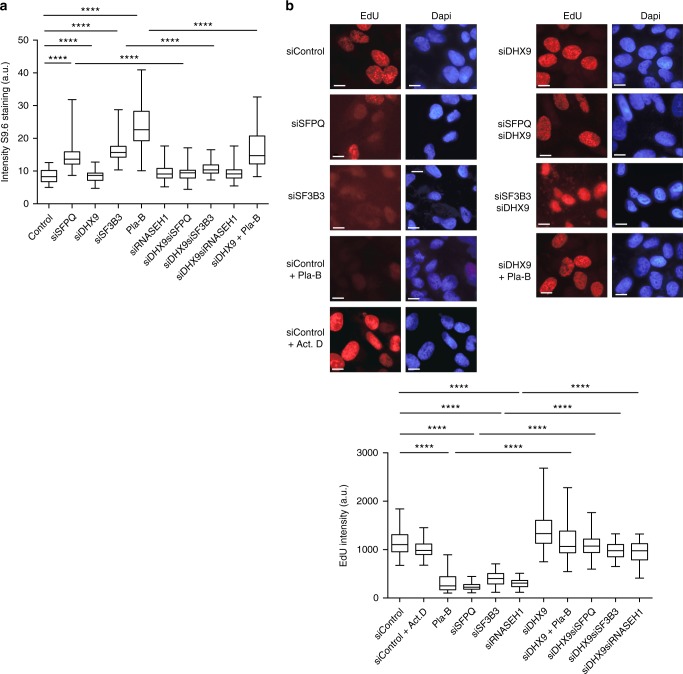
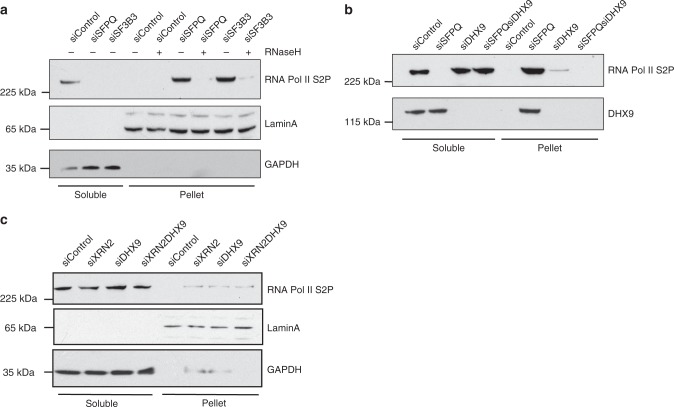
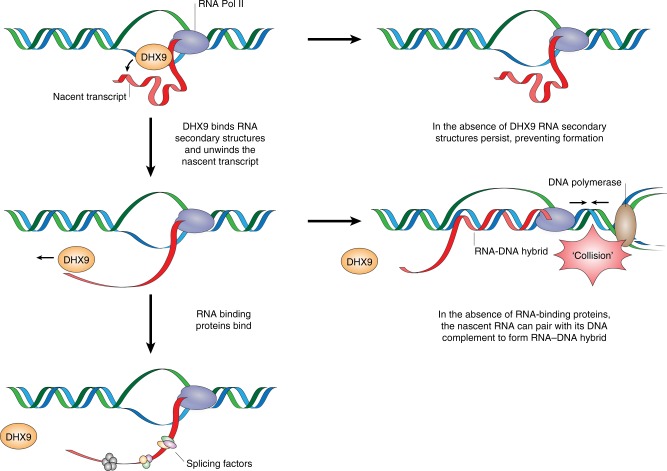
Similar articles
-
Human DHX9 helicase preferentially unwinds RNA-containing displacement loops (R-loops) and G-quadruplexes.DNA Repair (Amst). 2011 Jun 10;10(6):654-65. doi: 10.1016/j.dnarep.2011.04.013. Epub 2011 May 10. DNA Repair (Amst). 2011. PMID: 21561811
-
Complex interplay between FMRP and DHX9 during DNA replication stress.J Biol Chem. 2024 Jan;300(1):105572. doi: 10.1016/j.jbc.2023.105572. Epub 2023 Dec 16. J Biol Chem. 2024. PMID: 38110032 Free PMC article.
-
TDRD3 promotes DHX9 chromatin recruitment and R-loop resolution.Nucleic Acids Res. 2021 Sep 7;49(15):8573-8591. doi: 10.1093/nar/gkab642. Nucleic Acids Res. 2021. PMID: 34329467 Free PMC article.
-
The biology of DHX9 and its potential as a therapeutic target.Oncotarget. 2016 Jul 5;7(27):42716-42739. doi: 10.18632/oncotarget.8446. Oncotarget. 2016. PMID: 27034008 Free PMC article. Review.
-
R loops: new modulators of genome dynamics and function.Nat Rev Genet. 2015 Oct;16(10):583-97. doi: 10.1038/nrg3961. Epub 2015 Sep 15. Nat Rev Genet. 2015. PMID: 26370899 Review.
Cited by
-
Human Survival Motor Neuron genes generate a vast repertoire of circular RNAs.Nucleic Acids Res. 2019 Apr 8;47(6):2884-2905. doi: 10.1093/nar/gkz034. Nucleic Acids Res. 2019. PMID: 30698797 Free PMC article.
-
R-loop and its functions at the regulatory interfaces between transcription and (epi)genome.Biochim Biophys Acta Gene Regul Mech. 2021 Nov-Dec;1864(11-12):194750. doi: 10.1016/j.bbagrm.2021.194750. Epub 2021 Aug 28. Biochim Biophys Acta Gene Regul Mech. 2021. PMID: 34461314 Free PMC article. Review.
-
Locus-specific transcription silencing at the FHIT gene suppresses replication stress-induced copy number variant formation and associated replication delay.Nucleic Acids Res. 2021 Jul 21;49(13):7507-7524. doi: 10.1093/nar/gkab559. Nucleic Acids Res. 2021. PMID: 34181717 Free PMC article.
-
Looping forward: exploring R-loop processing and therapeutic potential.FEBS Lett. 2025 Jan;599(2):244-266. doi: 10.1002/1873-3468.14947. Epub 2024 Jun 6. FEBS Lett. 2025. PMID: 38844597 Free PMC article. Review.
-
Resolution of R-loops by topoisomerase III-β (TOP3B) in coordination with the DEAD-box helicase DDX5.Cell Rep. 2022 Jul 12;40(2):111067. doi: 10.1016/j.celrep.2022.111067. Cell Rep. 2022. PMID: 35830799 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
