

Host lipid sensing promotes invasion of cells with pathogenic Salmonella
- PMID: 30341337
- PMCID: PMC6195605
- DOI: 10.1038/s41598-018-33319-9
Host lipid sensing promotes invasion of cells with pathogenic Salmonella
Abstract
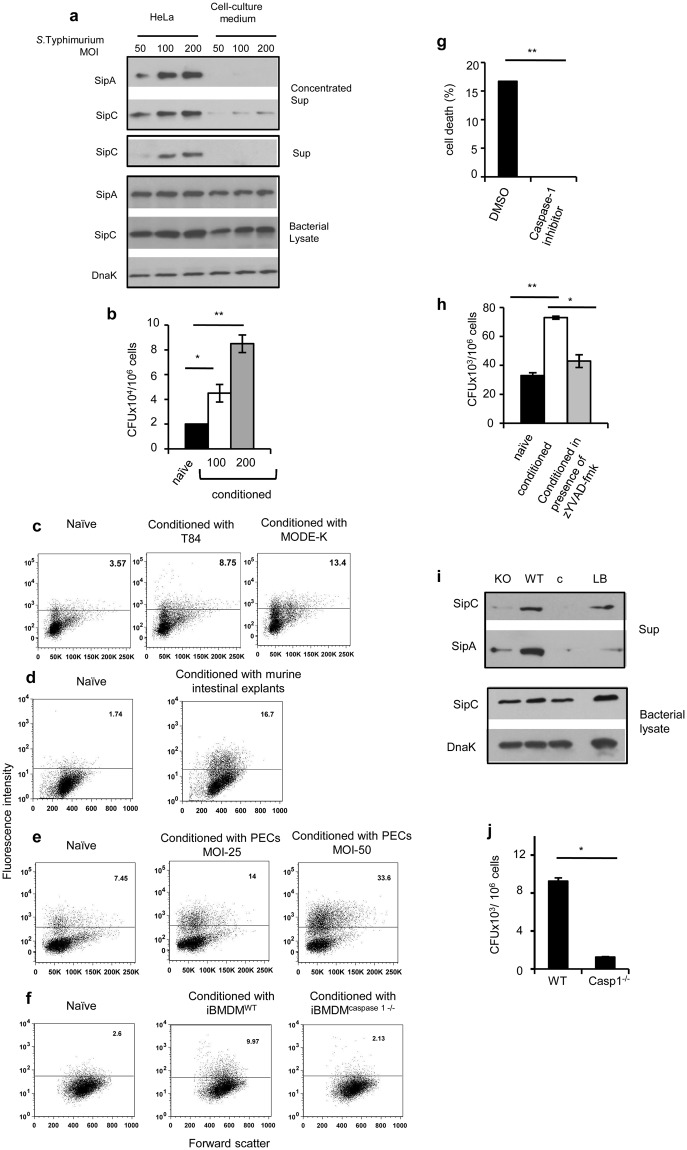
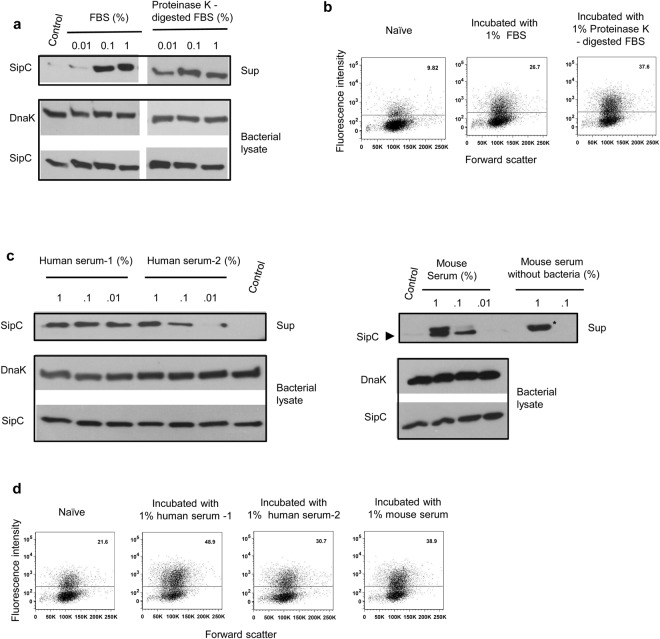
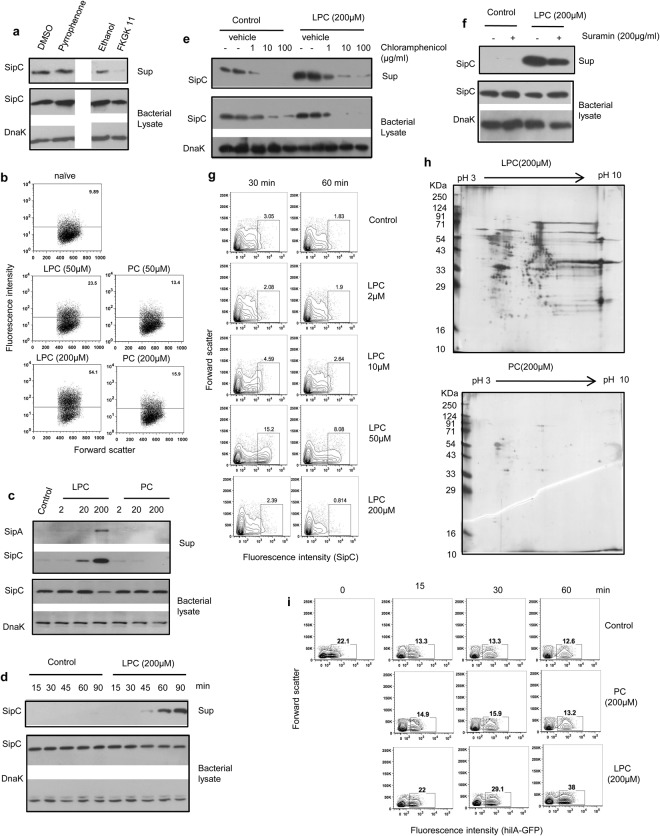
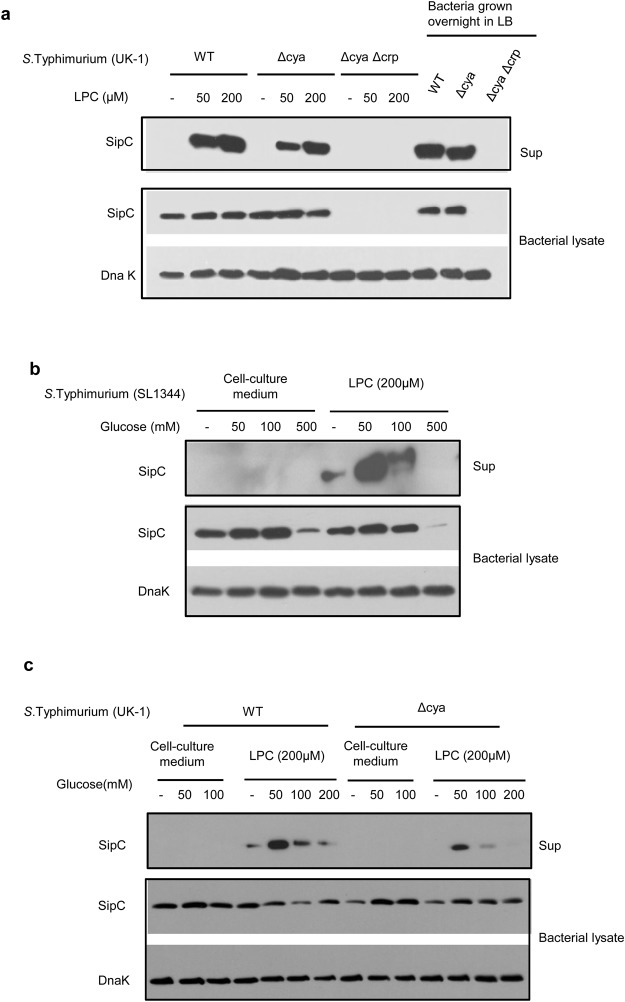
Pathogenic Salmonella species initiate infection by invading non-phagocytic intestinal epithelial cells (IEC). This invasion is brought about by a number of Salmonella invasion promoting molecules (Sips) encoded by the Salmonella Pathogenicity Island - 1 (SPI-1). Intracellular delivery of some of these molecules also brings about caspase-1 - mediated pyroptotic cell death that contributes to pathogen clearance. These molecules are secreted and delivered inside cells upon contact of Salmonella with one or more host signals whose identity has not been established. We show that lysophosphatidylcholine (LPC) released following activation of caspase-1 in Salmonella - infected cells and abundant in plasma amplifies production of Sips from this pathogen and promotes its cellular invasion. LPC brings about adenylate cyclase and cAMP receptor protein (CRP) - dependent de novo synthesis of SipC that is accompanied by its translocation to bacterial cell surface and release into the outside milieu. Treatment of Salmonella with LPC produces sustained induction of SPI - 1 transcriptional regulator, hilA. Our findings reveal a novel host lipid sensing - driven regulatory mechanism for Salmonella invasion.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Salmonella enterica Serovar Typhimurium SPI-1 and SPI-2 Shape the Global Transcriptional Landscape in a Human Intestinal Organoid Model System.mBio. 2021 May 18;12(3):e00399-21. doi: 10.1128/mBio.00399-21. mBio. 2021. PMID: 34006652 Free PMC article.
-
Salmonella pathogenicity island 2-dependent macrophage death is mediated in part by the host cysteine protease caspase-1.Cell Microbiol. 2001 Dec;3(12):825-37. doi: 10.1046/j.1462-5822.2001.00162.x. Cell Microbiol. 2001. PMID: 11736994
-
CRP-cAMP mediates silencing of Salmonella virulence at the post-transcriptional level.PLoS Genet. 2018 Jun 7;14(6):e1007401. doi: 10.1371/journal.pgen.1007401. eCollection 2018 Jun. PLoS Genet. 2018. PMID: 29879120 Free PMC article.
-
Evolution of Salmonella-Host Cell Interactions through a Dynamic Bacterial Genome.Front Cell Infect Microbiol. 2017 Sep 29;7:428. doi: 10.3389/fcimb.2017.00428. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 29034217 Free PMC article. Review.
-
The role of host cell death in Salmonella infections.Curr Top Microbiol Immunol. 2005;289:131-50. doi: 10.1007/3-540-27320-4_6. Curr Top Microbiol Immunol. 2005. PMID: 15791954 Review.
Cited by
-
Lysophosphatidylcholine Potentiates Antibacterial Activity of Polymyxin B.Antimicrob Agents Chemother. 2020 Nov 17;64(12):e01337-20. doi: 10.1128/AAC.01337-20. Print 2020 Nov 17. Antimicrob Agents Chemother. 2020. PMID: 32988824 Free PMC article.
-
The Role of Egg Yolk in Modulating the Virulence of Salmonella Enterica Serovar Enteritidis.Front Cell Infect Microbiol. 2022 Jun 14;12:903979. doi: 10.3389/fcimb.2022.903979. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35774398 Free PMC article.
-
SirA, CsrBC and HilD form in vivo a regulatory cascade that controls the SP1-1 and SPI-2 gene expression when Salmonella Typhimurium is in the intestinal lumen and are required for cecal colonization and liver dissemination in the avian model.Arch Microbiol. 2025 Apr 2;207(5):108. doi: 10.1007/s00203-025-04305-3. Arch Microbiol. 2025. PMID: 40169403 Free PMC article.
-
Recent progress in molecular mechanisms of Salmonella effectors involved in gut epithelium invasion.Mol Biol Rep. 2025 Jun 16;52(1):601. doi: 10.1007/s11033-025-10715-9. Mol Biol Rep. 2025. PMID: 40522423 Review.
-
Salmonella Pathogenicity Island 1 (SPI-1) and Its Complex Regulatory Network.Front Cell Infect Microbiol. 2019 Jul 31;9:270. doi: 10.3389/fcimb.2019.00270. eCollection 2019. Front Cell Infect Microbiol. 2019. PMID: 31428589 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
