

Stimulus strength determines the BTK-dependence of the SHIP1-deficient phenotype in IgE/antigen-triggered mast cells
- PMID: 30341350
- PMCID: PMC6195619
- DOI: 10.1038/s41598-018-33769-1
Stimulus strength determines the BTK-dependence of the SHIP1-deficient phenotype in IgE/antigen-triggered mast cells
Abstract
Antigen (Ag)-mediated crosslinking of IgE-loaded high-affinity receptors for IgE (FcεRI) on mast cells (MCs) triggers activation of proinflammatory effector functions relevant for IgE-associated allergic disorders. The cytosolic tyrosine kinase BTK and the SH2-containing inositol-5'-phosphatase SHIP1 are central positive and negative regulators of Ag-triggered MC activation, respectively, contrarily controlling Ca2+ mobilisation, degranulation, and cytokine production. Using genetic and pharmacological techniques, we examined whether BTK activation in Ship1-/- MCs is mandatory for the manifestation of the well-known hyperactive phenotype of Ship1-/- MCs. We demonstrate the prominence of BTK for the Ship1-/- phenotype in a manner strictly dependent on the strength of the initial Ag stimulus; particular importance for BTK was identified in Ship1-/- bone marrow-derived MCs in response to stimulation with suboptimal Ag concentrations. With respect to MAPK activation, BTK showed particular importance at suboptimal Ag concentrations, allowing for an analogous-to-digital switch resulting in full activation of ERK1/2 already at low Ag concentrations. Our data allow for a more precise definition of the role of BTK in FcεRI-mediated signal transduction and effector function in MCs. Moreover, they suggest that reduced activation or curtate expression of SHIP1 can be compensated by pharmacological inhibition of BTK and vice versa.
Conflict of interest statement
The authors declare no competing interests.
Figures
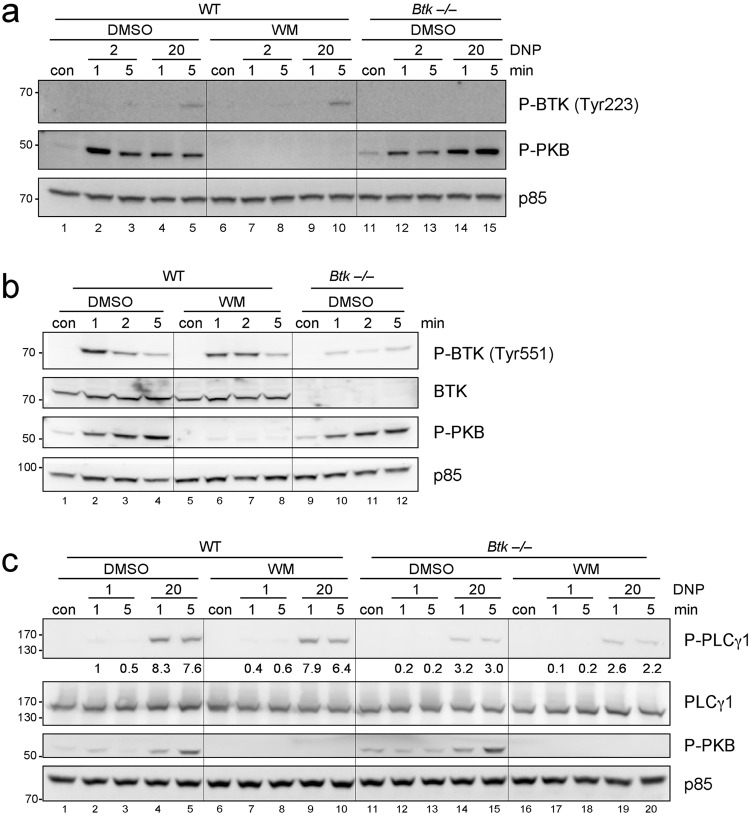
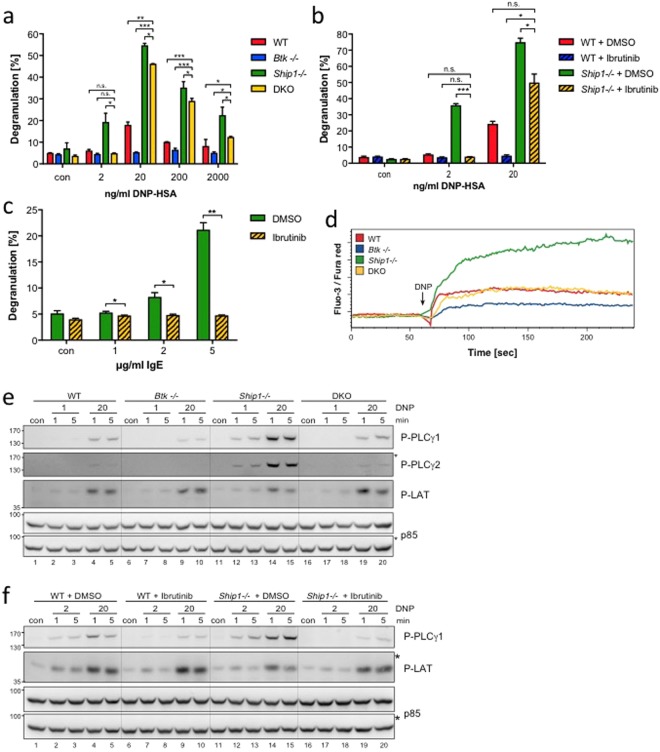
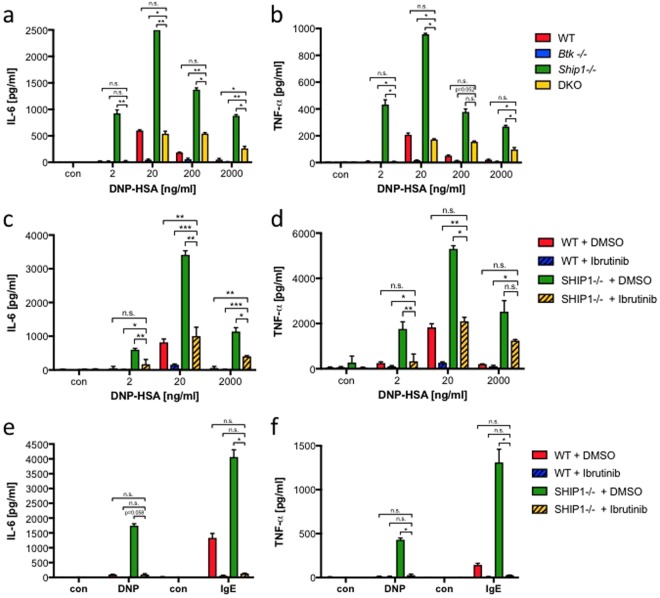
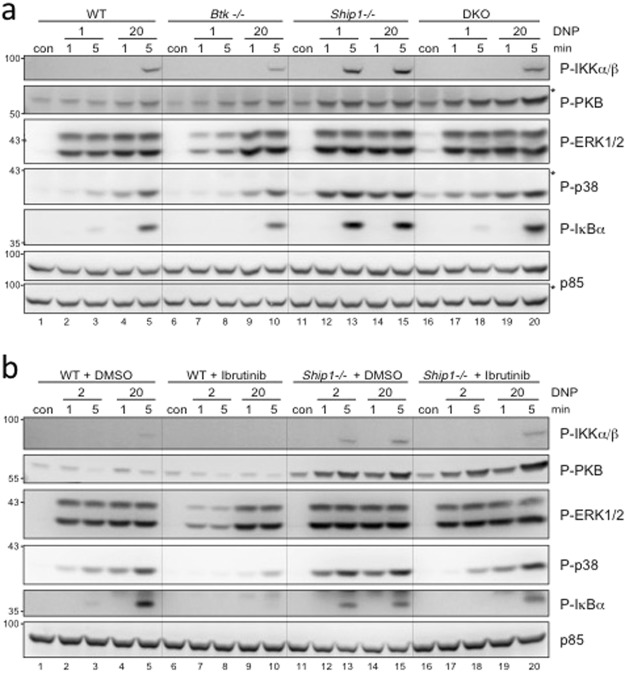
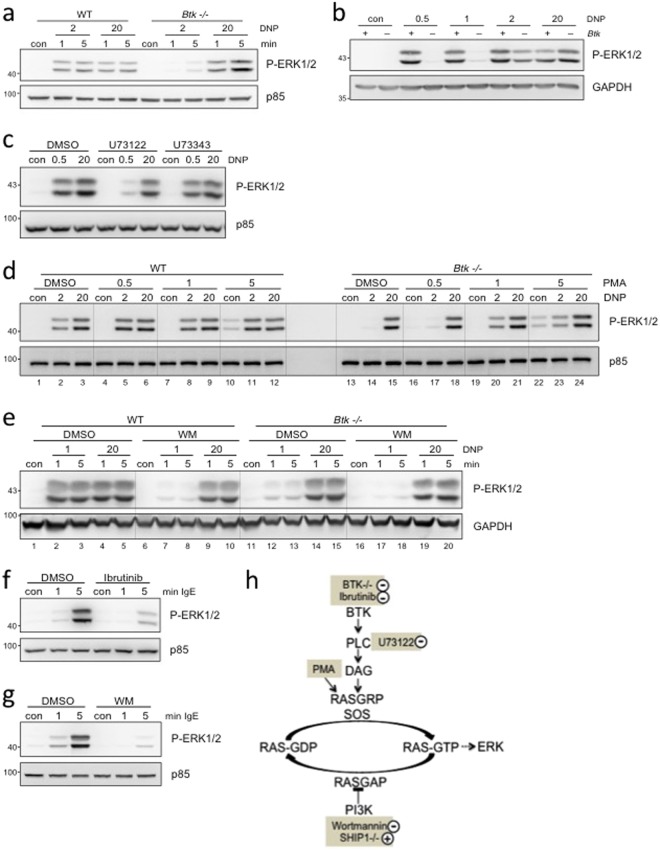
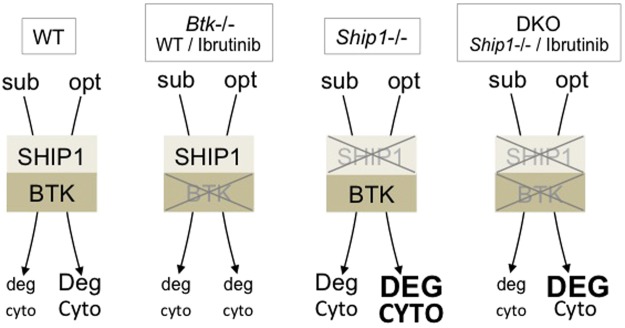
Similar articles
-
Differential Lyn-dependence of the SHIP1-deficient mast cell phenotype.Cell Commun Signal. 2016 May 20;14(1):12. doi: 10.1186/s12964-016-0135-0. Cell Commun Signal. 2016. PMID: 27206658 Free PMC article.
-
Differential use of BTK and PLC in FcεRI- and KIT-mediated mast cell activation: A marginal role of BTK upon KIT activation.Biochim Biophys Acta Mol Cell Res. 2020 Apr;1867(4):118622. doi: 10.1016/j.bbamcr.2019.118622. Epub 2019 Dec 11. Biochim Biophys Acta Mol Cell Res. 2020. PMID: 31837347
-
IgG-Independent Co-aggregation of FcεRI and FcγRIIB Results in LYN- and SHIP1-Dependent Tyrosine Phosphorylation of FcγRIIB in Murine Bone Marrow-Derived Mast Cells.Front Immunol. 2018 Aug 27;9:1937. doi: 10.3389/fimmu.2018.01937. eCollection 2018. Front Immunol. 2018. PMID: 30210494 Free PMC article.
-
Regulation of FcεRI signaling by lipid phosphatases.Curr Top Microbiol Immunol. 2014;382:111-27. doi: 10.1007/978-3-319-07911-0_6. Curr Top Microbiol Immunol. 2014. PMID: 25116098 Review.
-
SHIP1 and the negative control of mast cell/basophil activation by supra-optimal antigen concentrations.Mol Immunol. 2015 Jan;63(1):32-7. doi: 10.1016/j.molimm.2014.02.017. Epub 2014 Mar 25. Mol Immunol. 2015. PMID: 24679713 Review.
Cited by
-
Enzyme Is the Name-Adapter Is the Game.Cells. 2024 Jul 25;13(15):1249. doi: 10.3390/cells13151249. Cells. 2024. PMID: 39120280 Free PMC article. Review.
-
The Immunomodulatory Mechanisms of BTK Inhibition in CLL and Beyond.Cancers (Basel). 2024 Oct 23;16(21):3574. doi: 10.3390/cancers16213574. Cancers (Basel). 2024. PMID: 39518015 Free PMC article. Review.
-
The role of Bruton's tyrosine kinase in the immune system and disease.Immunology. 2021 Dec;164(4):722-736. doi: 10.1111/imm.13416. Epub 2021 Oct 4. Immunology. 2021. PMID: 34534359 Free PMC article. Review.
-
Bruton tyrosine kinase (BTK) may be a potential therapeutic target for interstitial cystitis/bladder pain syndrome.Aging (Albany NY). 2022 Sep 5;14(17):7052-7064. doi: 10.18632/aging.204271. Epub 2022 Sep 5. Aging (Albany NY). 2022. PMID: 36069808 Free PMC article.
-
Cytoskeletal Transport, Reorganization, and Fusion Regulation in Mast Cell-Stimulus Secretion Coupling.Front Cell Dev Biol. 2021 Mar 16;9:652077. doi: 10.3389/fcell.2021.652077. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33796537 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
