

Shared and genetically distinct Zea mays transcriptome responses to ongoing and past low temperature exposure
- PMID: 30342485
- PMCID: PMC6196024
- DOI: 10.1186/s12864-018-5134-7
Shared and genetically distinct Zea mays transcriptome responses to ongoing and past low temperature exposure
Abstract
Background: Cold temperatures and their alleviation affect many plant traits including the abundance of protein coding gene transcripts. Transcript level changes that occur in response to cold temperatures and their alleviation are shared or vary across genotypes. In this study we identify individual transcripts and groups of functionally related transcripts that consistently respond to cold and its alleviation. Genes that respond differently to temperature changes across genotypes may have limited functional importance. We investigate if these genes share functions, and if their genotype-specific gene expression levels change in magnitude or rank across temperatures.
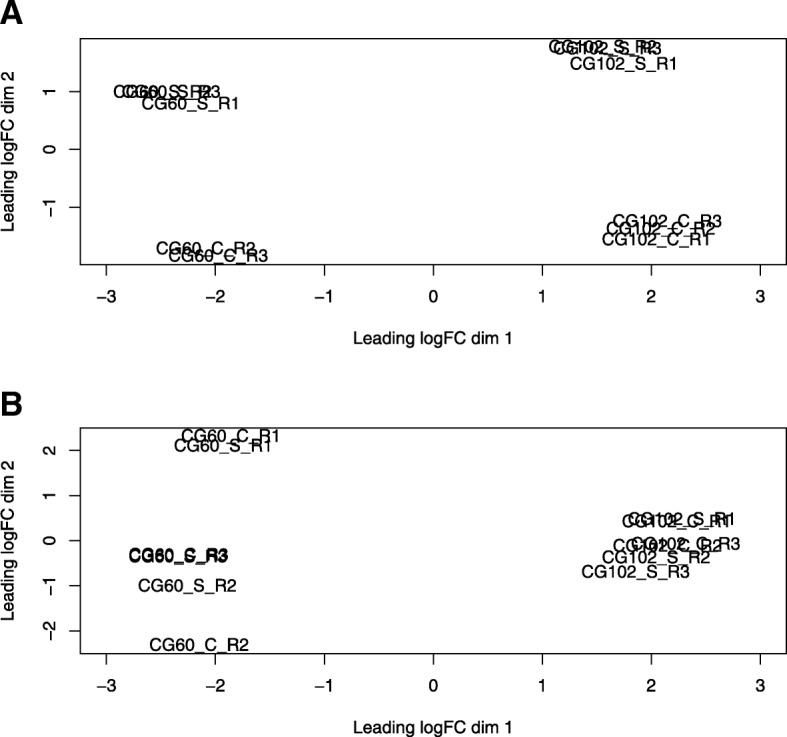
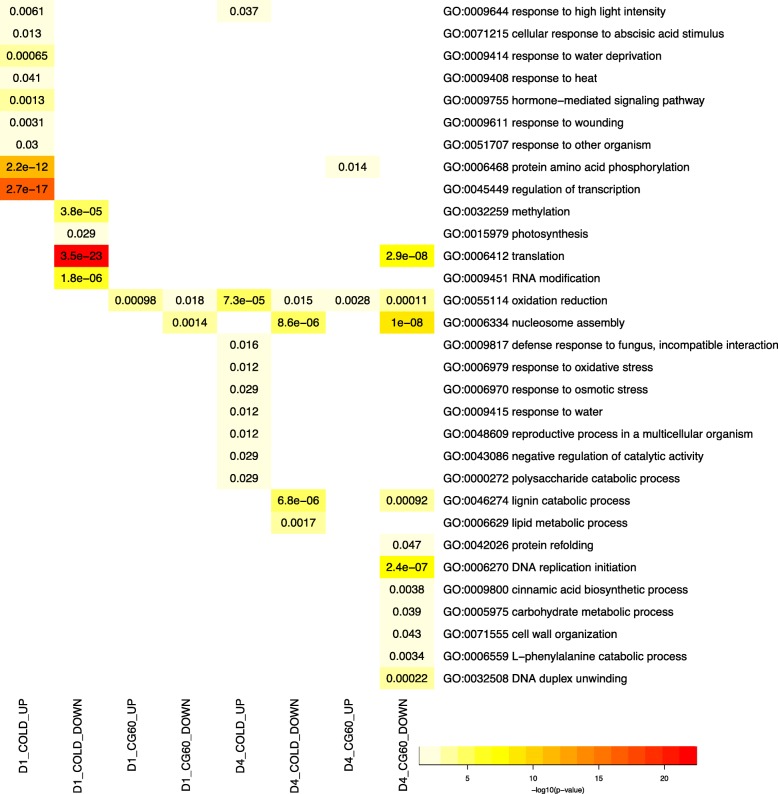
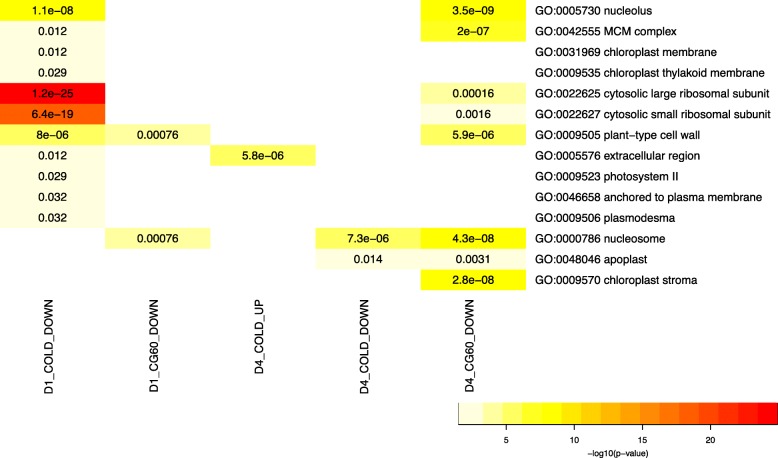
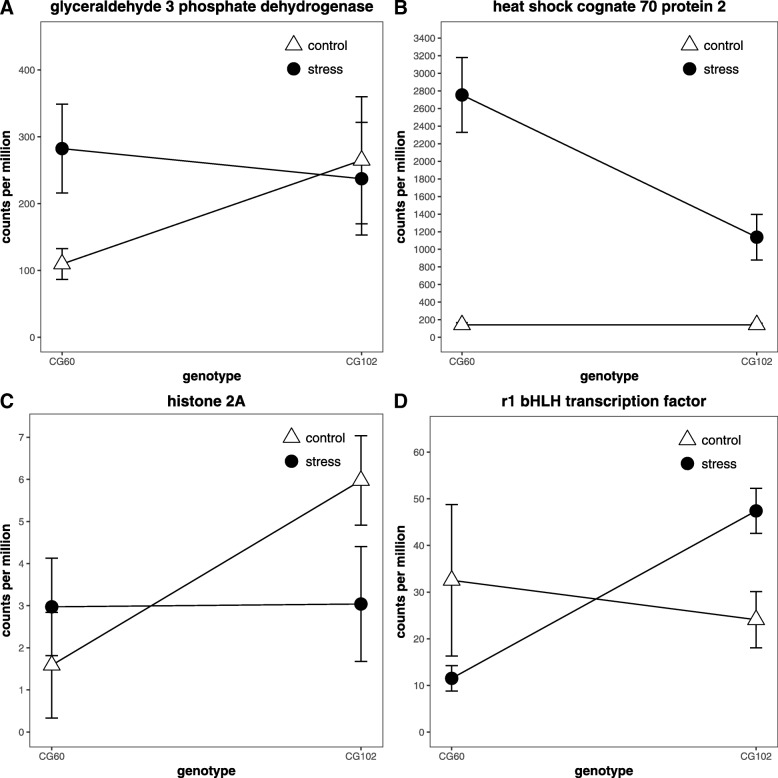
Results: We estimate transcript abundances from over 22,000 genes in two unrelated Zea mays inbred lines during and after cold temperature exposure. Genotype and temperature contribute to many genes' abundances. Past cold exposure affects many fewer genes. Genes up-regulated in cold encode many cytokinin glucoside biosynthesis enzymes, transcription factors, signalling molecules, and proteins involved in diverse environmental responses. After cold exposure, protease inhibitors and cuticular wax genes are newly up-regulated, and environmentally responsive genes continue to be up-regulated. Genes down-regulated in response to cold include many photosynthesis, translation, and DNA replication associated genes. After cold exposure, DNA replication and translation genes are still preferentially downregulated. Lignin and suberin biosynthesis are newly down-regulated. DNA replication, reactive oxygen species response, and anthocyanin biosynthesis genes have strong, genotype-specific temperature responses. The ranks of genotypes' transcript abundances often change across temperatures.
Conclusions: We report a large, core transcriptome response to cold and the alleviation of cold. In cold, many of the core suite of genes are up or downregulated to control plant growth and photosynthesis and limit cellular damage. In recovery, core responses are in part to prepare for future stress. Functionally related genes are consistently and greatly up-regulated in a single genotype in response to cold or its alleviation, suggesting positive selection has driven genotype-specific temperature responses in maize.
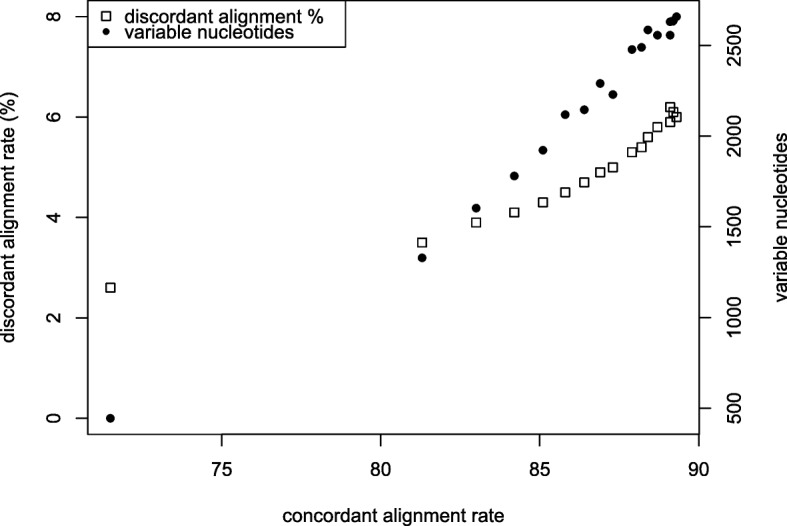
Keywords: Abiotic stress; Cold; Crossover interactions; Genotype environment interaction; Maize; RNA-Seq; Short read alignment.
Conflict of interest statement
Ethics approval and consent to participate
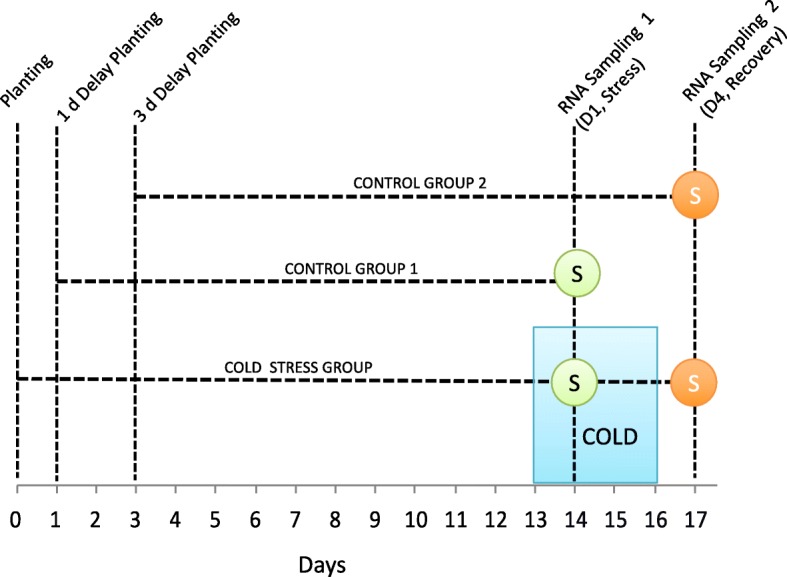
Maize inbred lines CG60 and CG102 were developed at the University of Guelph, and self-pollinated seed from these genotypes were generated in a Guelph, Ontario nursery. No field permissions were needed to generate the seed. Plants for this study were grown within controlled environments. The research complied with institutional, local, national, and international guidelines. No specimens have been deposited as vouchers.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Kocsy G, Pál M, Soltész A, Szalai G, Boldizsár Á, Kovács V, et al. Low temperature and oxidative stress in cereals. Acta Agron Hungarica. 2011;59:169–189. doi: 10.1556/AAgr.59.2011.2.7. - DOI
-
- Lee EA, Staebler MA, Tollenaar M. Genetic variation in physiological discriminators for cold tolerance — early autotrophic phase of development. Crop Sci. 2002;42:1919–1929. doi: 10.2135/cropsci2002.1919. - DOI
-
- Aroca R, Tognoni F, Irigoyen JJ, Sánchez-Díaz M, Pardossi A. Different root low temperature response of two maize genotypes differing in chilling sensitivity. Plant Physiol Biochem. 2001;39:1067–1073. doi: 10.1016/s0981-9428(01)01335-3. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
