

Aldosterone-sensitive HSD2 neurons in mice
- PMID: 30343334
- PMCID: PMC6369013
- DOI: 10.1007/s00429-018-1778-y
Aldosterone-sensitive HSD2 neurons in mice
Abstract
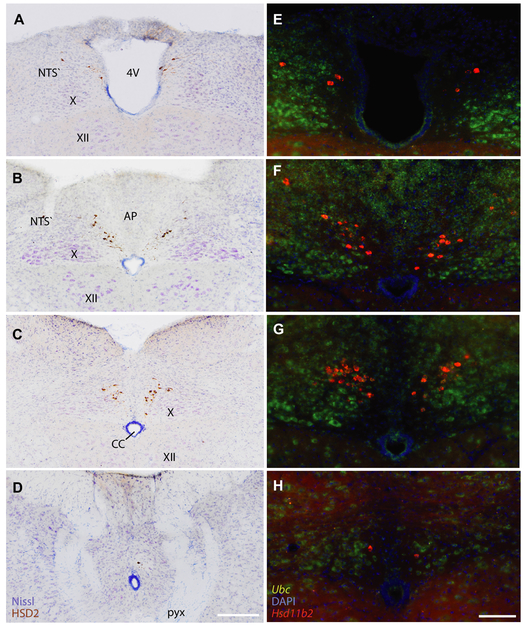
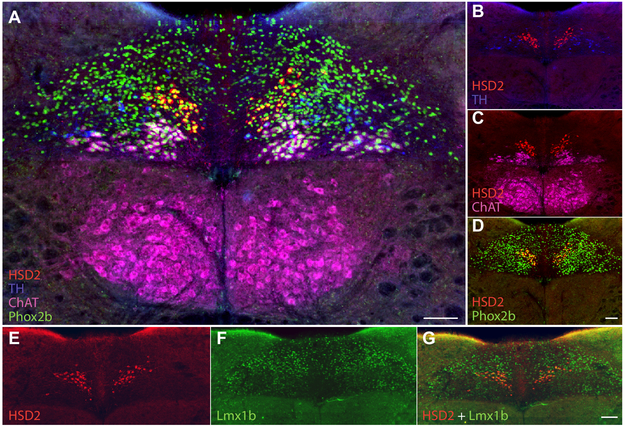
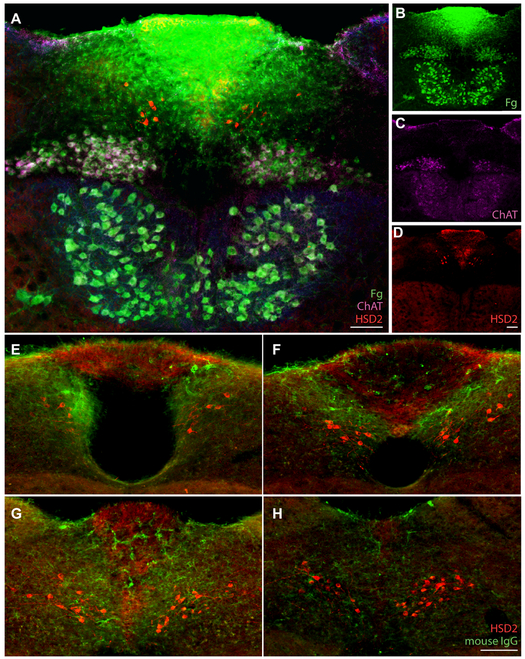
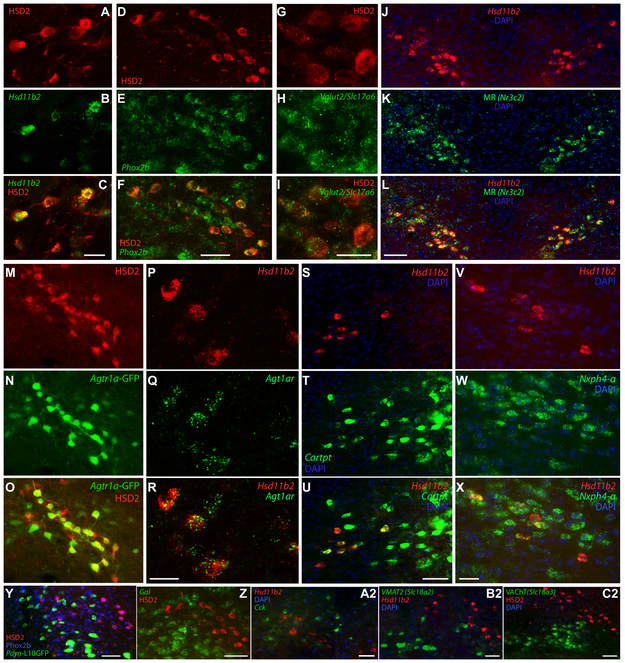
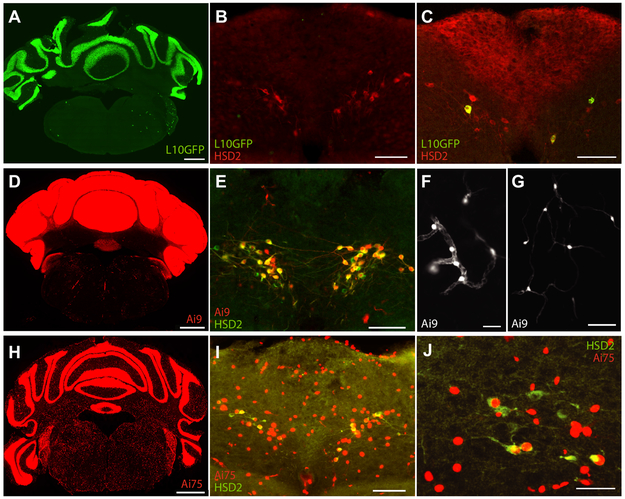
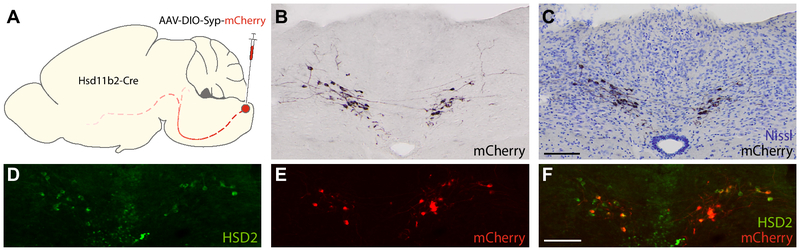
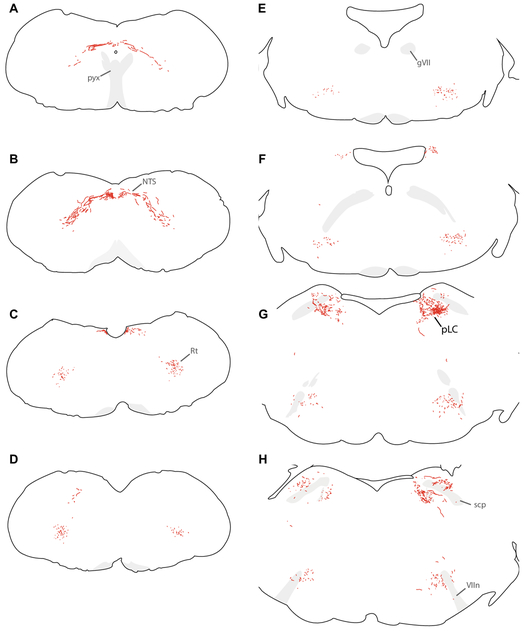
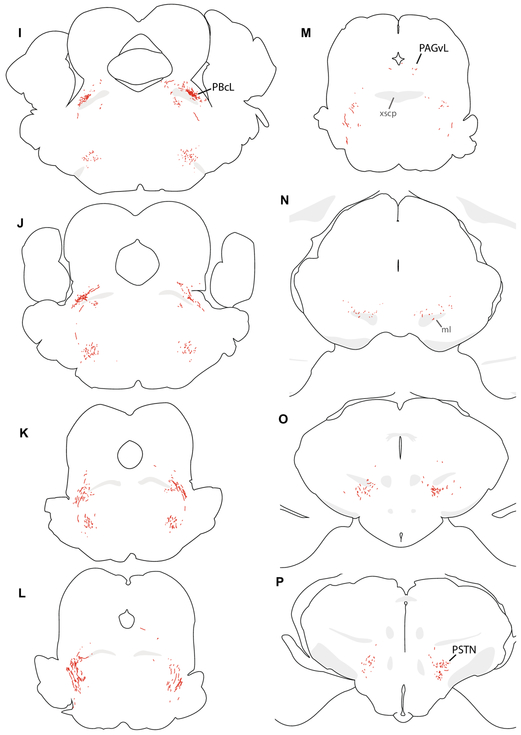
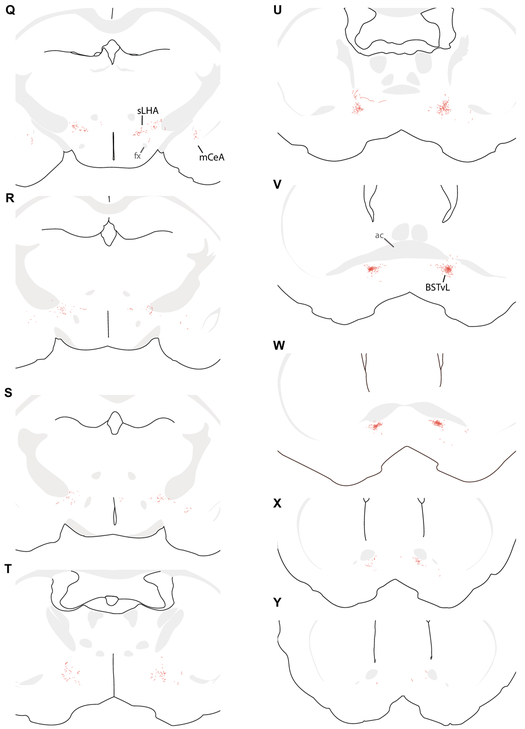
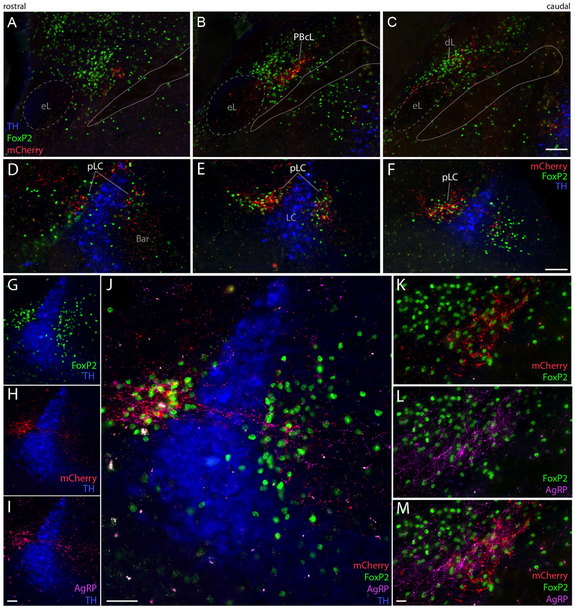
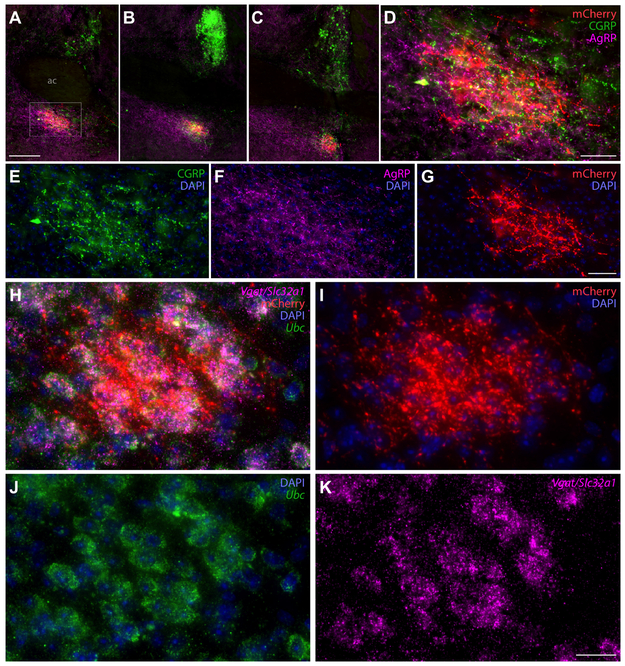
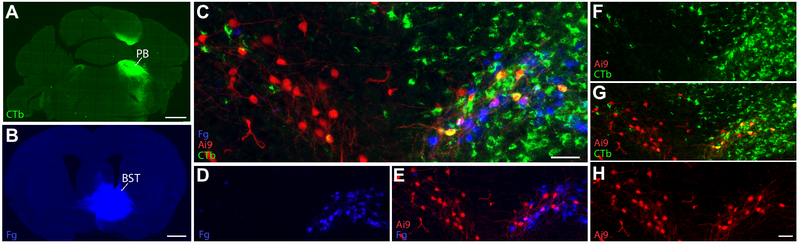
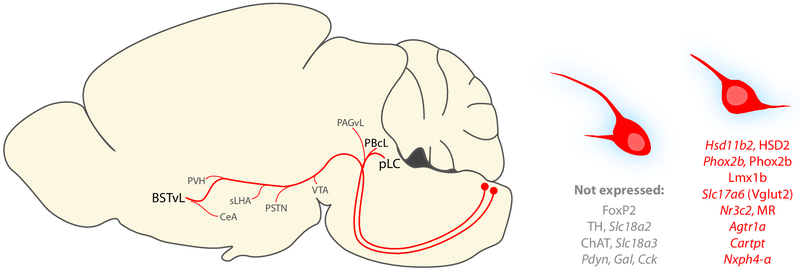
Sodium deficiency elevates aldosterone, which in addition to epithelial tissues acts on the brain to promote dysphoric symptoms and salt intake. Aldosterone boosts the activity of neurons that express 11-beta-hydroxysteroid dehydrogenase type 2 (HSD2), a hallmark of aldosterone-sensitive cells. To better characterize these neurons, we combine immunolabeling and in situ hybridization with fate mapping and Cre-conditional axon tracing in mice. Many cells throughout the brain have a developmental history of Hsd11b2 expression, but in the adult brain one small brainstem region with a leaky blood-brain barrier contains HSD2 neurons. These neurons express Hsd11b2, Nr3c2 (mineralocorticoid receptor), Agtr1a (angiotensin receptor), Slc17a6 (vesicular glutamate transporter 2), Phox2b, and Nxph4; many also express Cartpt or Lmx1b. No HSD2 neurons express cholinergic, monoaminergic, or several other neuropeptidergic markers. Their axons project to the parabrachial complex (PB), where they intermingle with AgRP-immunoreactive axons to form dense terminal fields overlapping FoxP2 neurons in the central lateral subnucleus (PBcL) and pre-locus coeruleus (pLC). Their axons also extend to the forebrain, intermingling with AgRP- and CGRP-immunoreactive axons to form dense terminals surrounding GABAergic neurons in the ventrolateral bed nucleus of the stria terminalis (BSTvL). Sparse axons target the periaqueductal gray, ventral tegmental area, lateral hypothalamic area, paraventricular hypothalamic nucleus, and central nucleus of the amygdala. Dual retrograde tracing revealed that largely separate HSD2 neurons project to pLC/PB or BSTvL. This projection pattern raises the possibility that a subset of HSD2 neurons promotes the dysphoric, anorexic, and anhedonic symptoms of hyperaldosteronism via AgRP-inhibited relay neurons in PB.
Keywords: 11-Beta-hydroxysteroid dehydrogenase type 2; Aldosterone; Angiotensin II; Dietary sodium; Dietary sodium deficiency; Dietary sodium deprivation; Mineralocorticoid receptor; Nucleus of the solitary tract; Salt appetite; Sodium appetite.
Figures
References
-
- Arriza JL, Simerly RB, Swanson LW, Evans RM. 1988. The neuronal mineralocorticoid receptor as a mediator of glucocorticoid response. Neuron 1(9):887–900. - PubMed
-
- Aston-Jones G, Delfs JM, Druhan J, Zhu Y. 1999. The bed nucleus of the stria terminalis. A target site for noradrenergic actions in opiate withdrawal. Ann N Y Acad Sci 877:486–498. - PubMed
-
- Bard P 1928. A diencephalic mechanism for the expression of rage with special reference to the sympathetic nervous system. American Journal of Physiology 84(3):490–515.
MeSH terms
Substances
Grants and funding
- F32 DK103387/National Institute of Diabetes and Digestive and Kidney Diseases
- NS099425/National Institute of Neurological Disorders and Stroke (US)
- K08 NS099425/NS/NINDS NIH HHS/United States
- JG2017/Aging Mind and Brain Institute, University of Iowa Center on Aging
- F32 DK103387/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
