

Ocular Clocks: Adapting Mechanisms for Eye Functions and Health
- PMID: 30347082
- PMCID: PMC6181243
- DOI: 10.1167/iovs.18-24957
Ocular Clocks: Adapting Mechanisms for Eye Functions and Health
Abstract
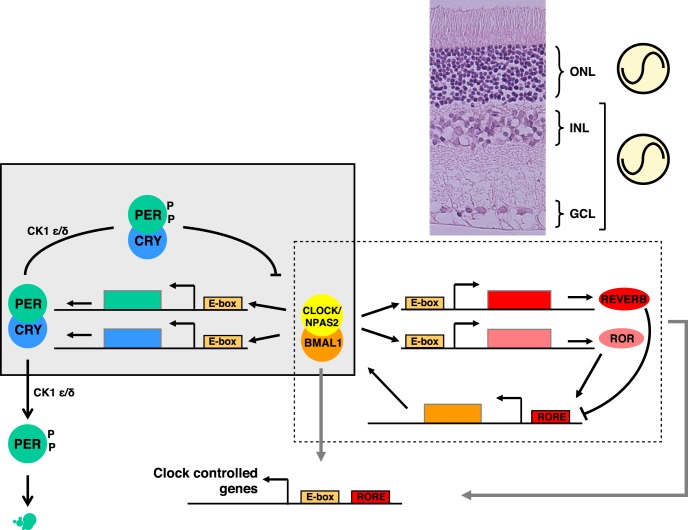
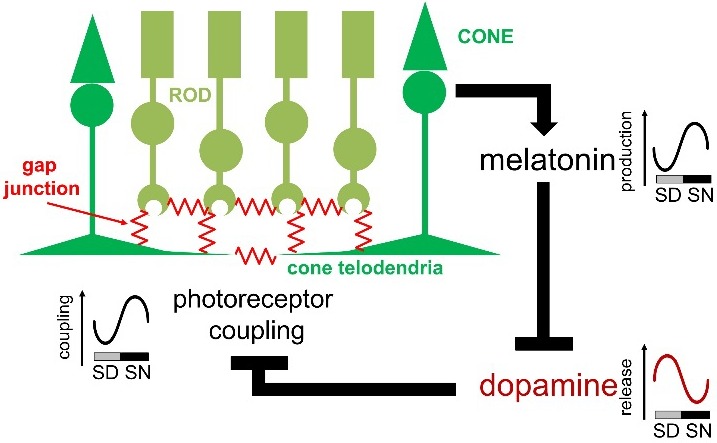
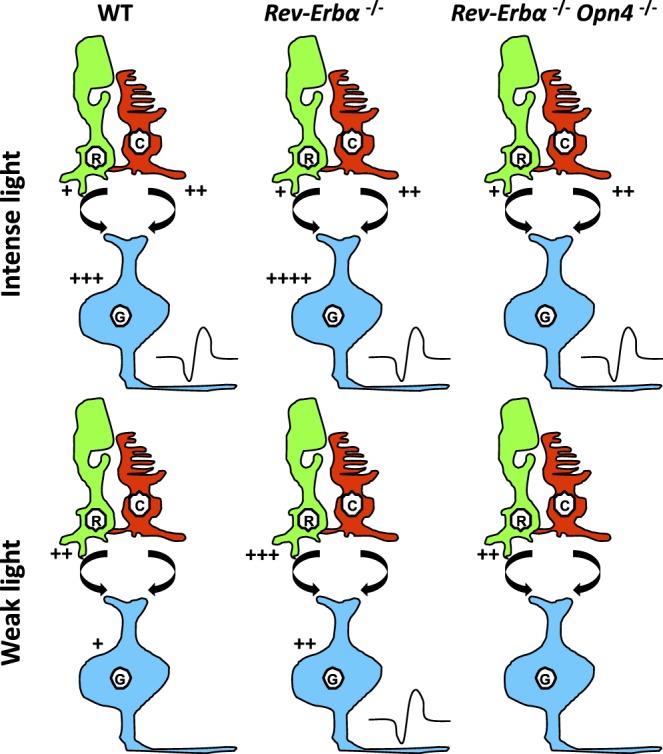
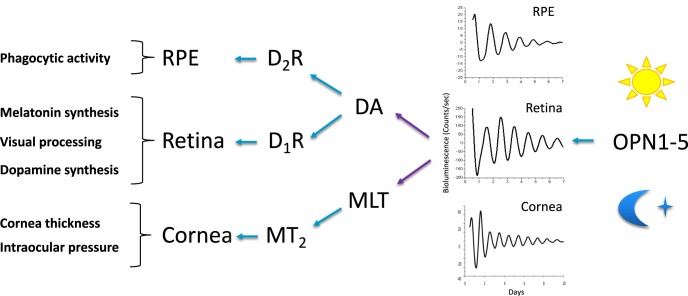
Vision is a highly rhythmic function adapted to the extensive changes in light intensity occurring over the 24-hour day. This adaptation relies on rhythms in cellular and molecular processes, which are orchestrated by a network of circadian clocks located within the retina and in the eye, synchronized to the day/night cycle and which, together, fine-tune detection and processing of light information over the 24-hour period and ensure retinal homeostasis. Systematic or high throughput studies revealed a series of genes rhythmically expressed in the retina, pointing at specific functions or pathways under circadian control. Conversely, knockout studies demonstrated that the circadian clock regulates retinal processing of light information. In addition, recent data revealed that it also plays a role in development as well as in aging of the retina. Regarding synchronization by the light/dark cycle, the retina displays the unique property of bringing together light sensitivity, clock machinery, and a wide range of rhythmic outputs. Melatonin and dopamine play a particular role in this system, being both outputs and inputs for clocks. The retinal cellular complexity suggests that mechanisms of regulation by light are diverse and intricate. In the context of the whole eye, the retina looks like a major determinant of phase resetting for other tissues such as the retinal pigmented epithelium or cornea. Understanding the pathways linking the cell-specific molecular machineries to their cognate outputs will be one of the major challenges for the future.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
