

Drosophila as a Model for Developmental Biology: Stem Cell-Fate Decisions in the Developing Nervous System
- PMID: 30347666
- PMCID: PMC6315890
- DOI: 10.3390/jdb6040025
Drosophila as a Model for Developmental Biology: Stem Cell-Fate Decisions in the Developing Nervous System
Abstract
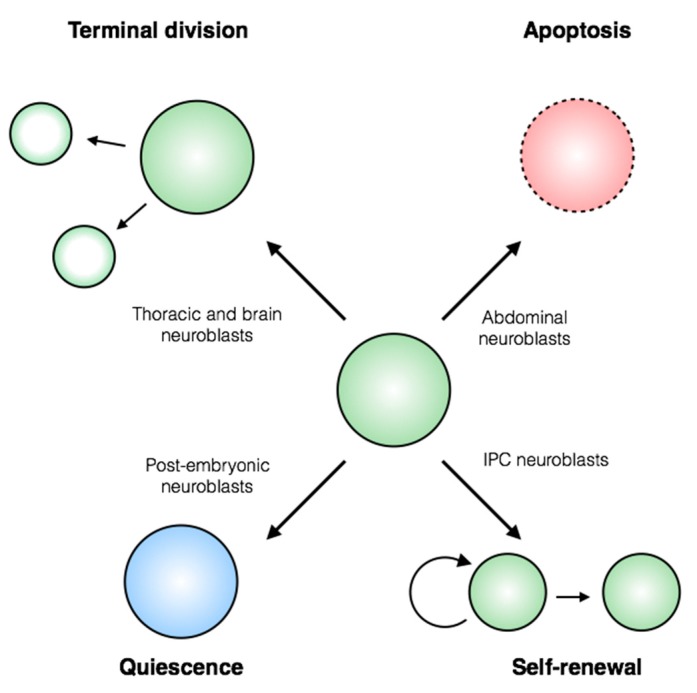
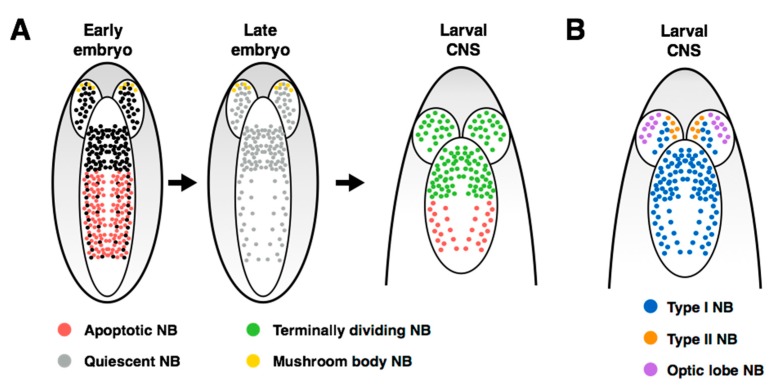
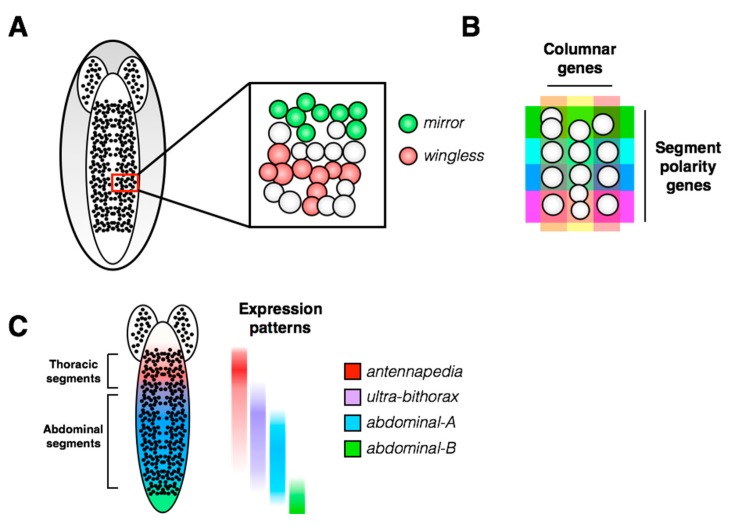
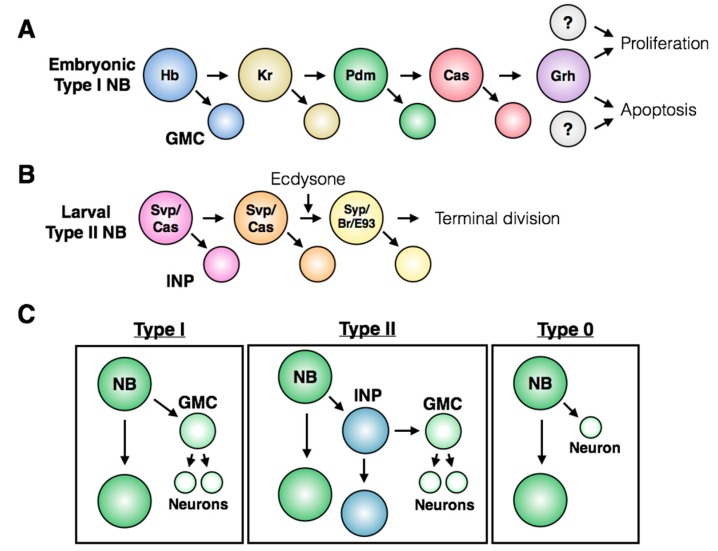
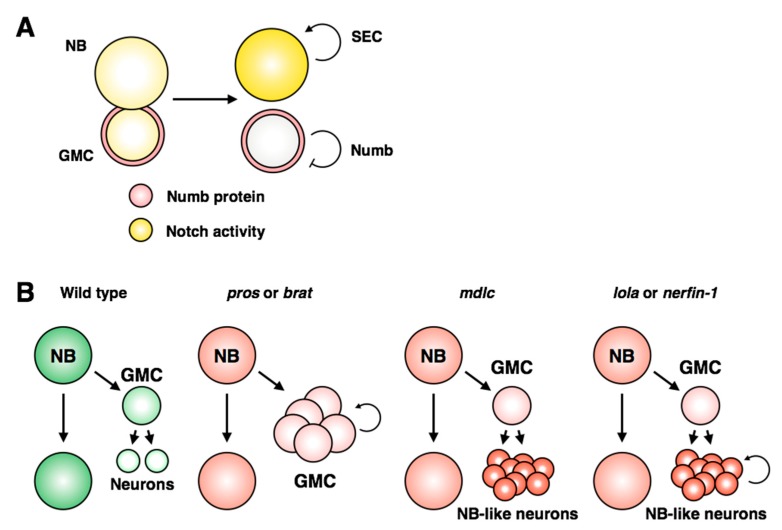
Stem cells face a diversity of choices throughout their lives. At specific times, they may decide to initiate cell division, terminal differentiation, or apoptosis, or they may enter a quiescent non-proliferative state. Neural stem cells in the Drosophila central nervous system do all of these, at stereotypical times and anatomical positions during development. Distinct populations of neural stem cells offer a unique system to investigate the regulation of a particular stem cell behavior, while comparisons between populations can lead us to a broader understanding of stem cell identity. Drosophila is a well-described and genetically tractable model for studying fundamental stem cell behavior and the mechanisms that underlie cell-fate decisions. This review will focus on recent advances in our understanding of the factors that contribute to distinct stem cell-fate decisions within the context of the Drosophila nervous system.
Keywords: Drosophila; apoptosis; cell cycle; development; neuroblast; stem cell biology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Decoupling developmental apoptosis and neuroblast proliferation in Drosophila.Dev Biol. 2019 Dec 1;456(1):17-24. doi: 10.1016/j.ydbio.2019.08.004. Epub 2019 Aug 4. Dev Biol. 2019. PMID: 31390535 Free PMC article.
-
Drosophila neural stem cells in brain development and tumor formation.J Neurogenet. 2014 Sep-Dec;28(3-4):181-9. doi: 10.3109/01677063.2014.898639. Epub 2014 May 12. J Neurogenet. 2014. PMID: 24766377 Review.
-
Drosophila neural stem cells: cell cycle control of self-renewal, differentiation, and termination in brain development.Results Probl Cell Differ. 2011;53:529-46. doi: 10.1007/978-3-642-19065-0_21. Results Probl Cell Differ. 2011. PMID: 21630158 Review.
-
ASC proneural factors are necessary for chromatin remodeling during neuroectodermal to neuroblast fate transition to ensure the timely initiation of the neural stem cell program.BMC Biol. 2022 May 13;20(1):107. doi: 10.1186/s12915-022-01300-8. BMC Biol. 2022. PMID: 35549704 Free PMC article.
-
Temporal control of Drosophila central nervous system development.Curr Opin Neurobiol. 2019 Jun;56:24-32. doi: 10.1016/j.conb.2018.10.016. Epub 2018 Nov 28. Curr Opin Neurobiol. 2019. PMID: 30500514 Review.
Cited by
-
Silver nanoparticle-induced developmental inhibition of Drosophila melanogaster accompanies disruption of genetic material of larval neural stem cells and non-neuronal cells.Environ Monit Assess. 2019 Jul 16;191(8):497. doi: 10.1007/s10661-019-7630-x. Environ Monit Assess. 2019. PMID: 31312907
-
Dedifferentiation-derived neural stem cells exhibit perturbed temporal progression.EMBO Rep. 2023 Jun 5;24(6):e55837. doi: 10.15252/embr.202255837. Epub 2023 Apr 11. EMBO Rep. 2023. PMID: 37039033 Free PMC article.
-
Non-autonomous regulation of neurogenesis by extrinsic cues: a Drosophila perspective.Oxf Open Neurosci. 2022 May 4;1:kvac004. doi: 10.1093/oons/kvac004. eCollection 2022. Oxf Open Neurosci. 2022. PMID: 38596708 Free PMC article. Review.
-
Mxc, a Drosophila homolog of mental retardation-associated gene NPAT, maintains neural stem cell fate.Cell Biosci. 2022 May 31;12(1):78. doi: 10.1186/s13578-022-00820-8. Cell Biosci. 2022. PMID: 35642004 Free PMC article.
-
Tools to reverse-engineer multicellular systems: case studies using the fruit fly.J Biol Eng. 2019 Apr 23;13:33. doi: 10.1186/s13036-019-0161-8. eCollection 2019. J Biol Eng. 2019. PMID: 31049075 Free PMC article. Review.
References
-
- Abrams J.M., White K., Fessier L.I., Steller H. Programmed cell death during Drosophila embryogenesis. Development. 1993;117:29–43. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
