

EHD2 is a mechanotransducer connecting caveolae dynamics with gene transcription
- PMID: 30348749
- PMCID: PMC6279385
- DOI: 10.1083/jcb.201801122
EHD2 is a mechanotransducer connecting caveolae dynamics with gene transcription
Abstract
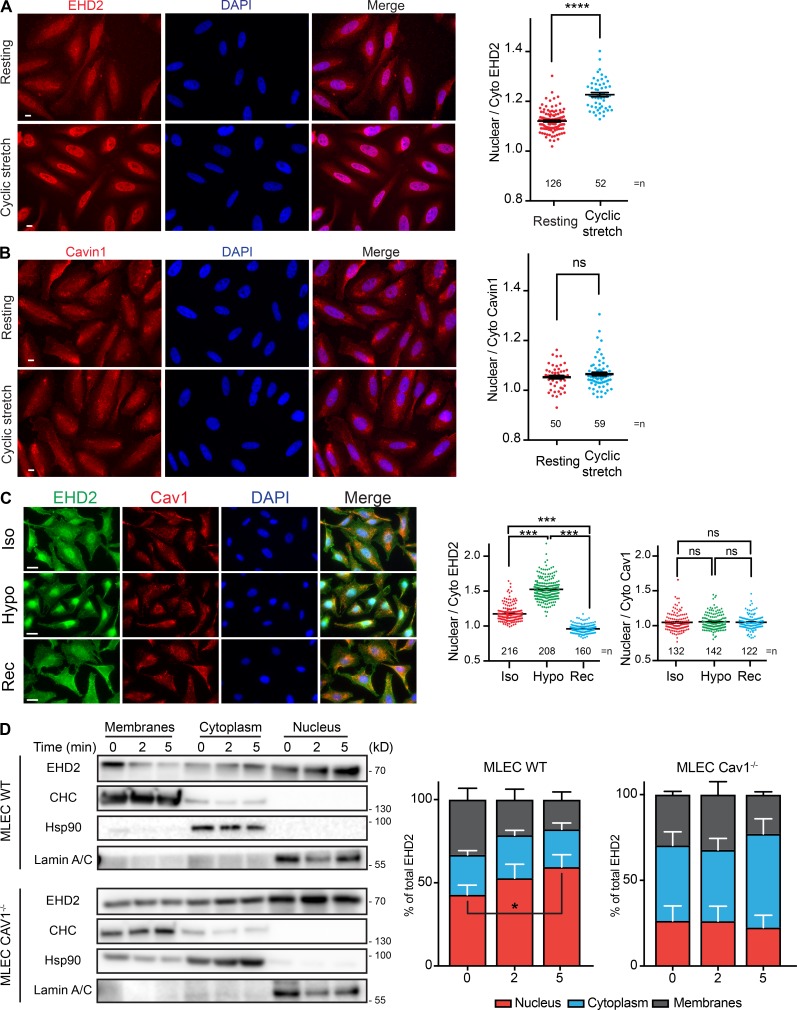
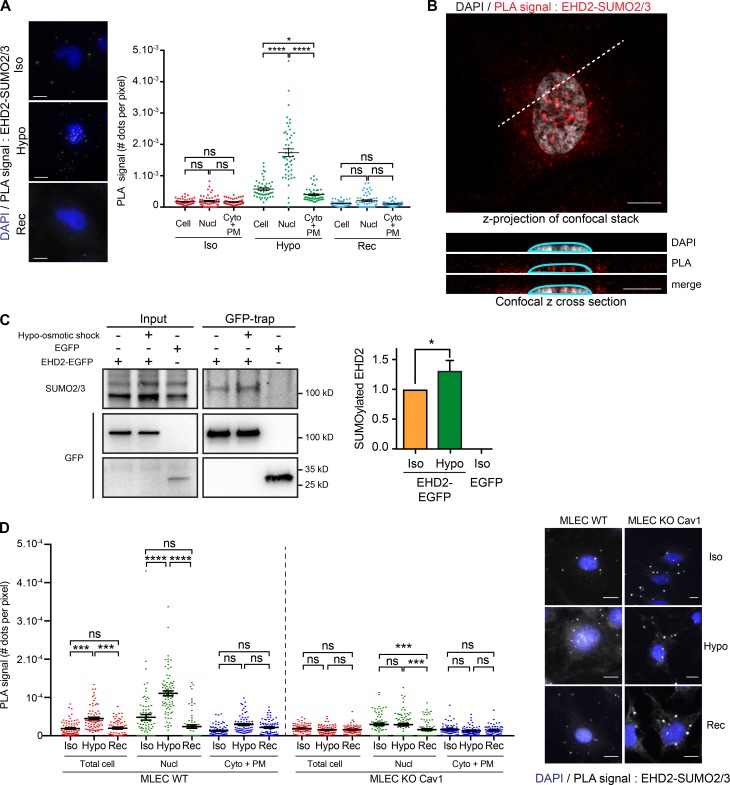
Caveolae are small invaginated pits that function as dynamic mechanosensors to buffer tension variations at the plasma membrane. Here we show that under mechanical stress, the EHD2 ATPase is rapidly released from caveolae, SUMOylated, and translocated to the nucleus, where it regulates the transcription of several genes including those coding for caveolae constituents. We also found that EHD2 is required to maintain the caveolae reservoir at the plasma membrane during the variations of membrane tension induced by mechanical stress. Metal-replica electron microscopy of breast cancer cells lacking EHD2 revealed a complete absence of caveolae and a lack of gene regulation under mechanical stress. Expressing EHD2 was sufficient to restore both functions in these cells. Our findings therefore define EHD2 as a central player in mechanotransduction connecting the disassembly of the caveolae reservoir with the regulation of gene transcription under mechanical stress.
© 2018 Torrino et al.
Figures



References
-
- Ariotti N., Fernández-Rojo M.A., Zhou Y., Hill M.M., Rodkey T.L., Inder K.L., Tanner L.B., Wenk M.R., Hancock J.F., and Parton R.G.. 2014. Caveolae regulate the nanoscale organization of the plasma membrane to remotely control Ras signaling. J. Cell Biol. 204:777–792. 10.1083/jcb.201307055 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
