

Structural and mechanistic insights into the function of the unconventional class XIV myosin MyoA from Toxoplasma gondii
- PMID: 30348763
- PMCID: PMC6233092
- DOI: 10.1073/pnas.1811167115
Structural and mechanistic insights into the function of the unconventional class XIV myosin MyoA from Toxoplasma gondii
Abstract
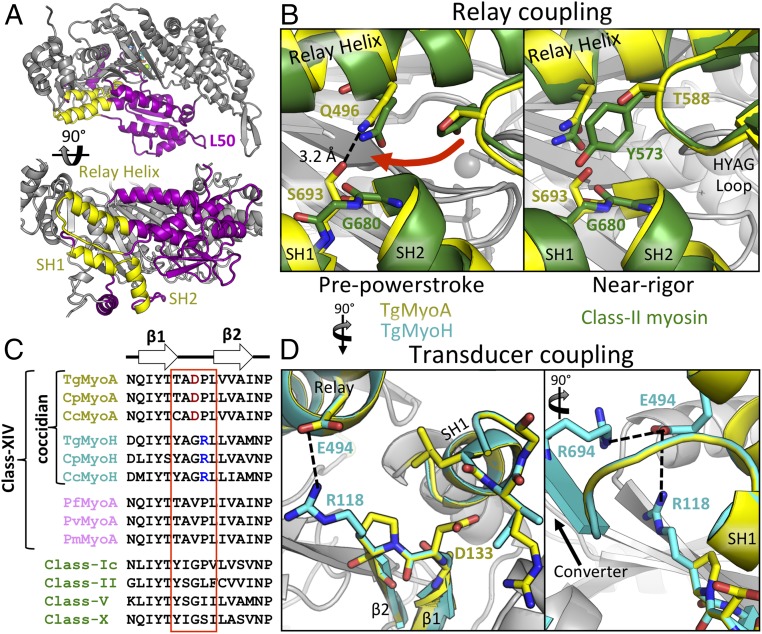
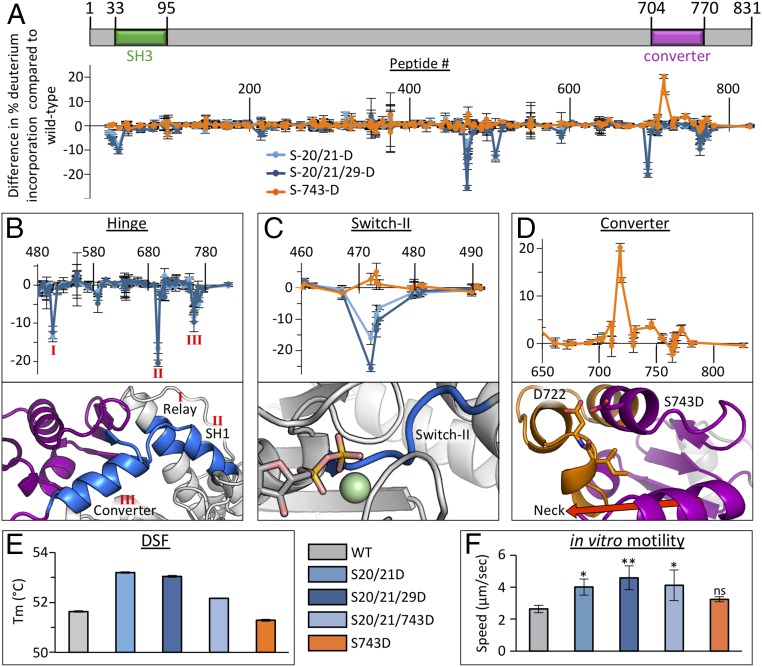
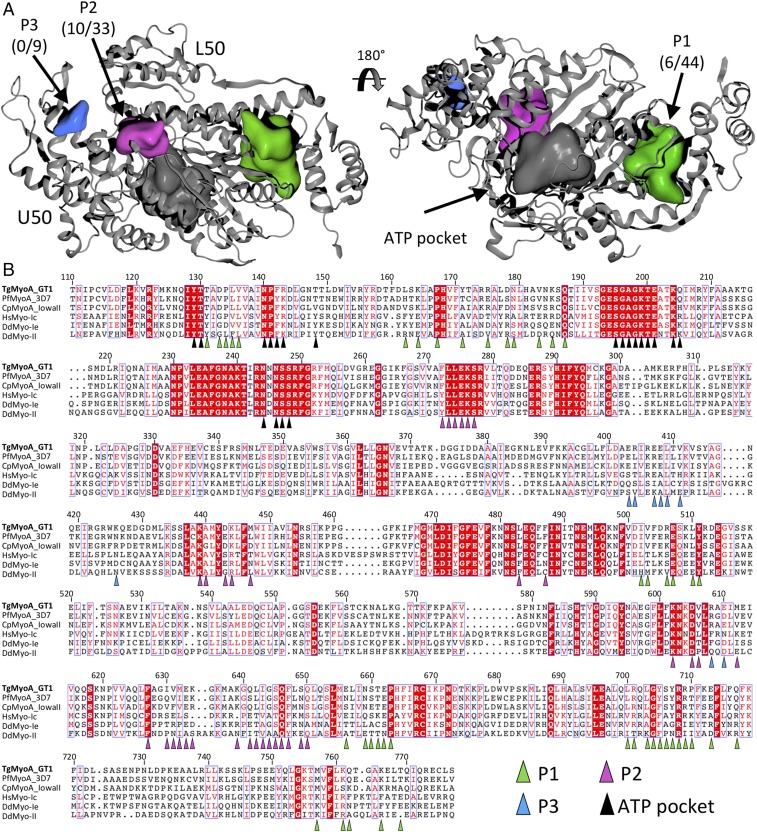
Parasites of the phylum Apicomplexa are responsible for significant morbidity and mortality on a global scale. Central to the virulence of these pathogens are the phylum-specific, unconventional class XIV myosins that power the essential processes of parasite motility and host cell invasion. Notably, class XIV myosins differ from human myosins in key functional regions, yet they are capable of fast movement along actin filaments with kinetics rivaling previously studied myosins. Toward establishing a detailed molecular mechanism of class XIV motility, we determined the 2.6-Å resolution crystal structure of the Toxoplasma gondii MyoA (TgMyoA) motor domain. Structural analysis reveals intriguing strategies for force transduction and chemomechanical coupling that rely on a divergent SH1/SH2 region, the class-defining "HYAG"-site polymorphism, and the actin-binding surface. In vitro motility assays and hydrogen-deuterium exchange coupled with MS further reveal the mechanistic underpinnings of phosphorylation-dependent modulation of TgMyoA motility whereby localized regions of increased stability and order correlate with enhanced motility. Analysis of solvent-accessible pockets reveals striking differences between apicomplexan class XIV and human myosins. Extending these analyses to high-confidence homology models of Plasmodium and Cryptosporidium MyoA motor domains supports the intriguing potential of designing class-specific, yet broadly active, apicomplexan myosin inhibitors. The successful expression of the functional TgMyoA complex combined with our crystal structure of the motor domain provides a strong foundation in support of detailed structure-function studies and enables the development of small-molecule inhibitors targeting these devastating global pathogens.
Keywords: Apicomplexa; Toxoplasma gondii; X-ray crystallography; motility; myosin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Harker KS, Ueno N, Lodoen MB. Toxoplasma gondii dissemination: A parasite’s journey through the infected host. Parasite Immunol. 2015;37:141–149. - PubMed
-
- Frenal K, Dubremetz JF, Lebrun M, Soldati-Favre D. Gliding motility powers invasion and egress in Apicomplexa. Nat Rev Microbiol. 2017;15:645–660. - PubMed
-
- Frenal K, Foth BJ, Soldati-Favre D. Myosin class XIV and other myosins inprotists. In: Coluccio LM, editor. Myosins: A Superfamily of Molecular Motors, Proteins and Cell Regulation. Vol 7. Springer; Dordrecht, The Netherlands: 2008. pp. 421–440.
-
- Mueller C, Graindorge A, Soldati-Favre D. Functions of myosin motors tailored for parasitism. Curr Opin Microbiol. 2017;40:113–122. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
