

Isolation and characterization of NY-ESO-1-specific T cell receptors restricted on various MHC molecules
- PMID: 30348802
- PMCID: PMC6233129
- DOI: 10.1073/pnas.1810653115
Isolation and characterization of NY-ESO-1-specific T cell receptors restricted on various MHC molecules
Abstract
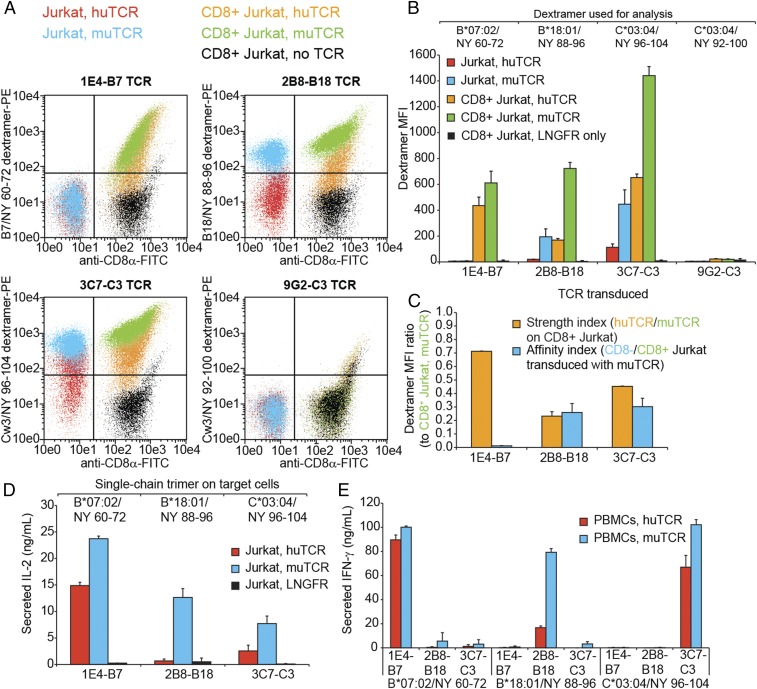
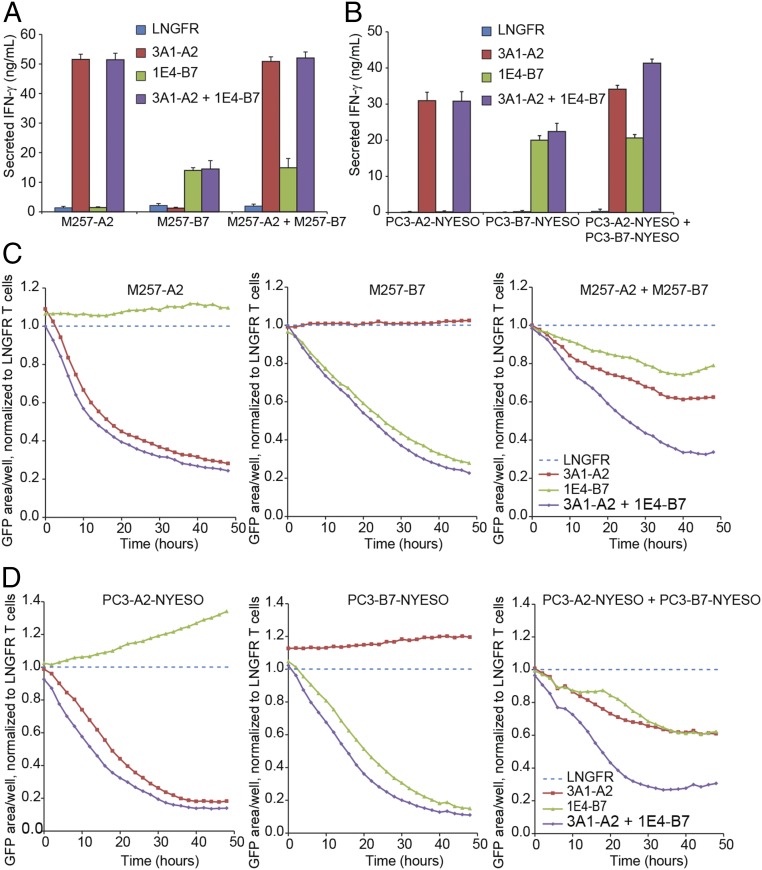
Tumor-specific T cell receptor (TCR) gene transfer enables specific and potent immune targeting of tumor antigens. Due to the prevalence of the HLA-A2 MHC class I supertype in most human populations, the majority of TCR gene therapy trials targeting public antigens have employed HLA-A2-restricted TCRs, limiting this approach to those patients expressing this allele. For these patients, TCR gene therapy trials have resulted in both tantalizing successes and lethal adverse events, underscoring the need for careful selection of antigenic targets. Broad and safe application of public antigen-targeted TCR gene therapies will require (i) selecting public antigens that are highly tumor-specific and (ii) targeting multiple epitopes derived from these antigens by obtaining an assortment of TCRs restricted by multiple common MHC alleles. The canonical cancer-testis antigen, NY-ESO-1, is not expressed in normal tissues but is aberrantly expressed across a broad array of cancer types. It has also been targeted with A2-restricted TCR gene therapy without adverse events or notable side effects. To enable the targeting of NY-ESO-1 in a broader array of HLA haplotypes, we isolated TCRs specific for NY-ESO-1 epitopes presented by four MHC molecules: HLA-A2, -B07, -B18, and -C03. Using these TCRs, we pilot an approach to extend TCR gene therapies targeting NY-ESO-1 to patient populations beyond those expressing HLA-A2.
Keywords: MHC; NY-ESO-1; T cell receptor gene therapy; TCR; immunotherapy.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
Conflict of interest statement: A patent application has been filed (serial no. 62/727,485) entitled “Composition of NY-ESO-1-Specific T Cell Receptors Restricted on Multiple Major Histocompatibility Complex Molecules.”
Figures
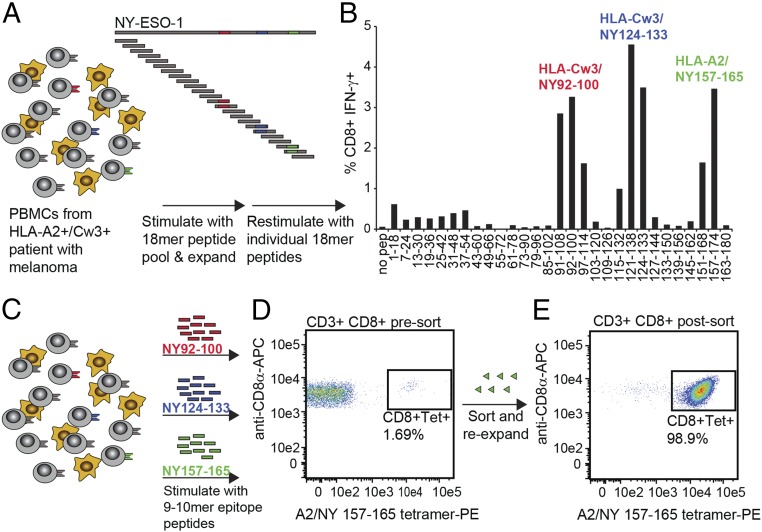
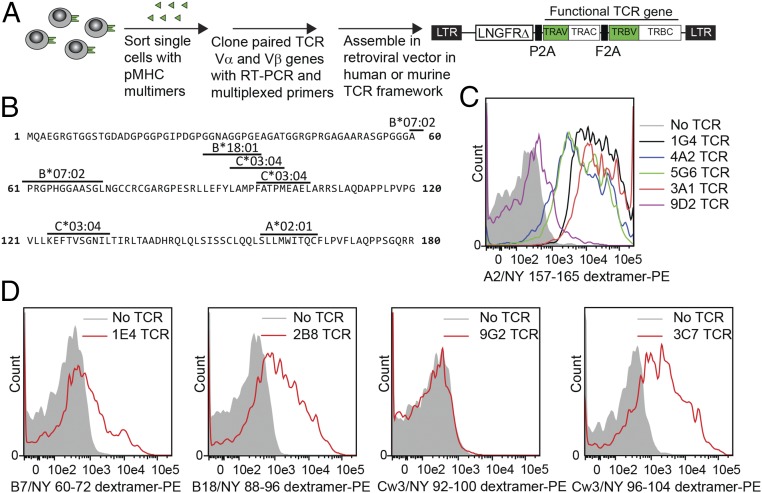
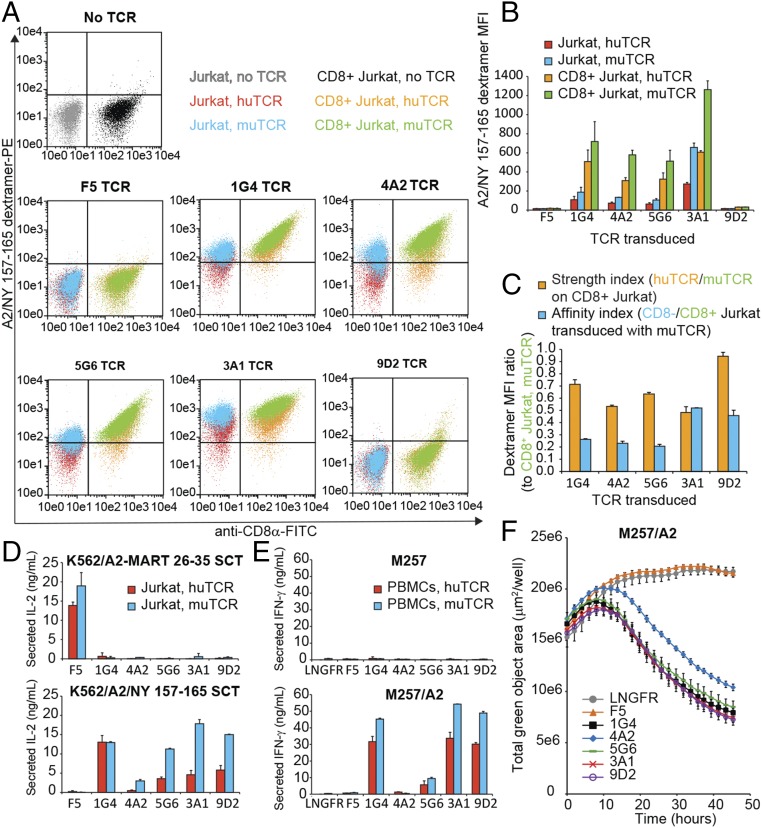
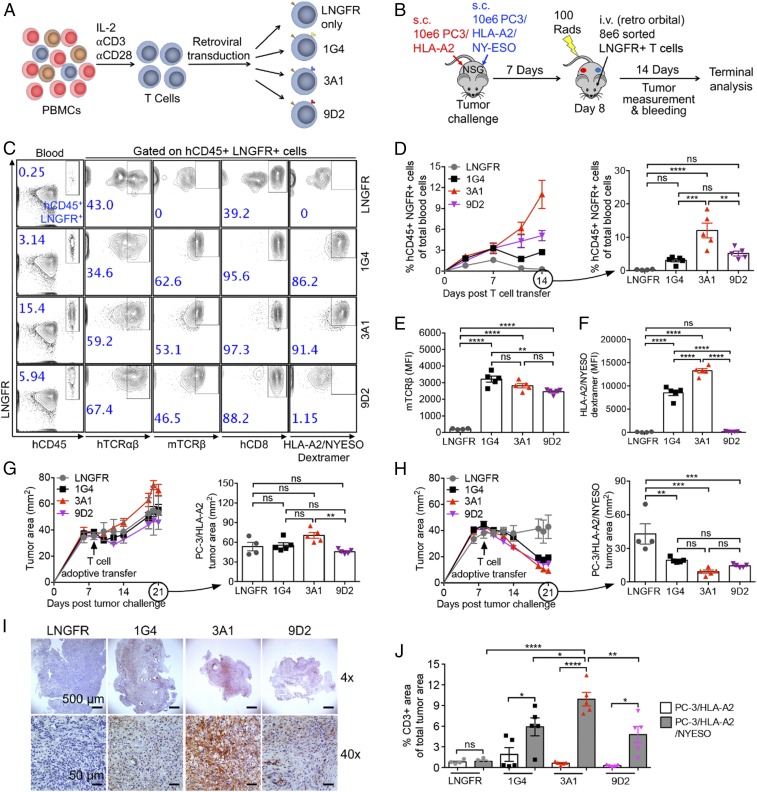
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
