

Essential Roles of the sppRA Fructose-Phosphate Phosphohydrolase Operon in Carbohydrate Metabolism and Virulence Expression by Streptococcus mutans
- PMID: 30348833
- PMCID: PMC6304665
- DOI: 10.1128/JB.00586-18
Essential Roles of the sppRA Fructose-Phosphate Phosphohydrolase Operon in Carbohydrate Metabolism and Virulence Expression by Streptococcus mutans
Abstract
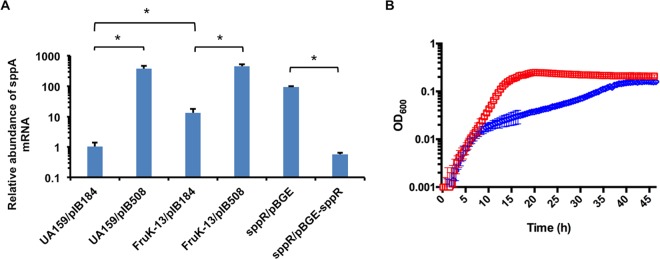
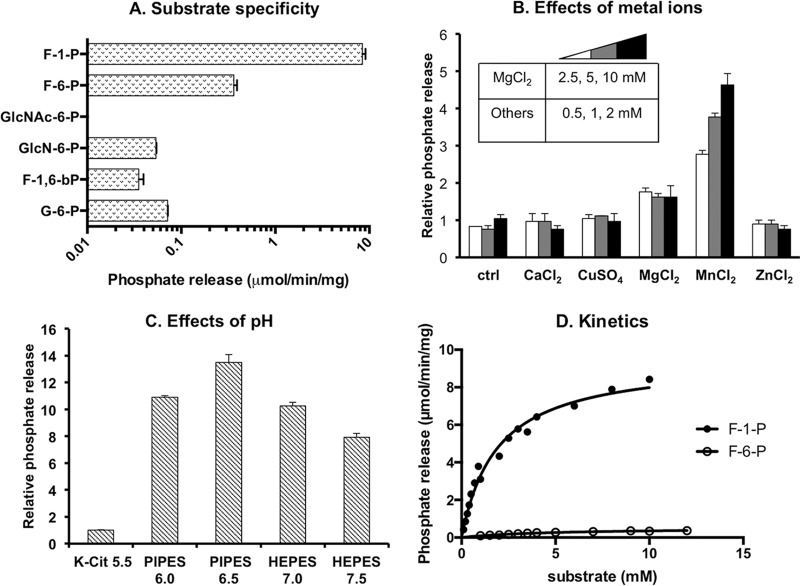
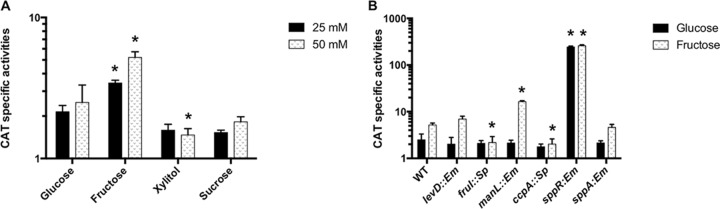
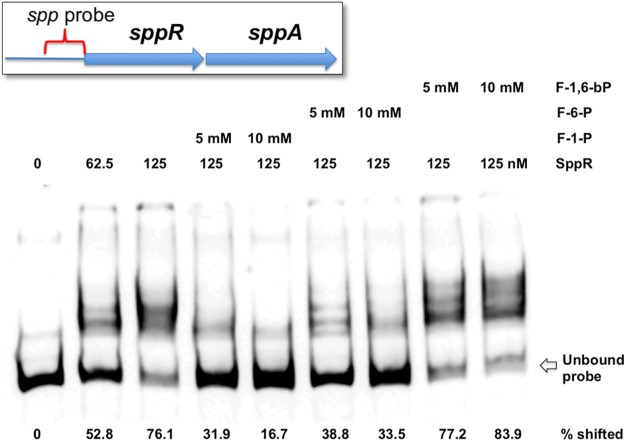
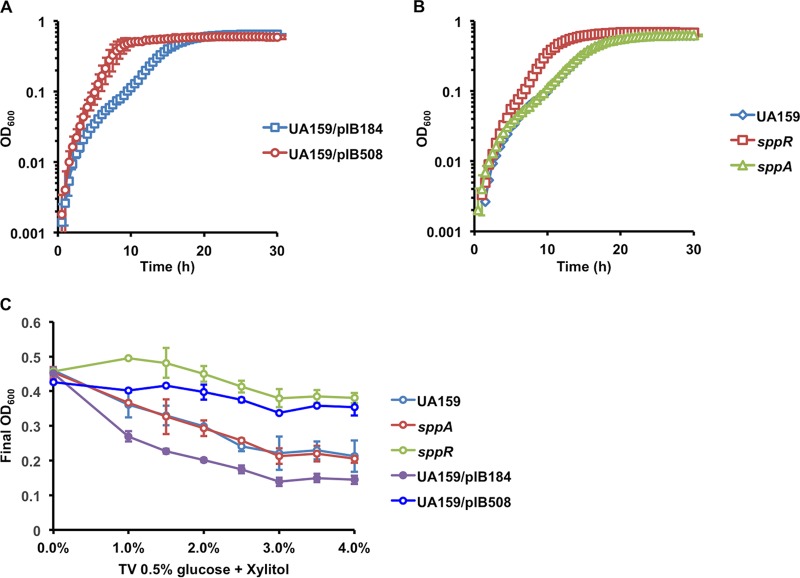
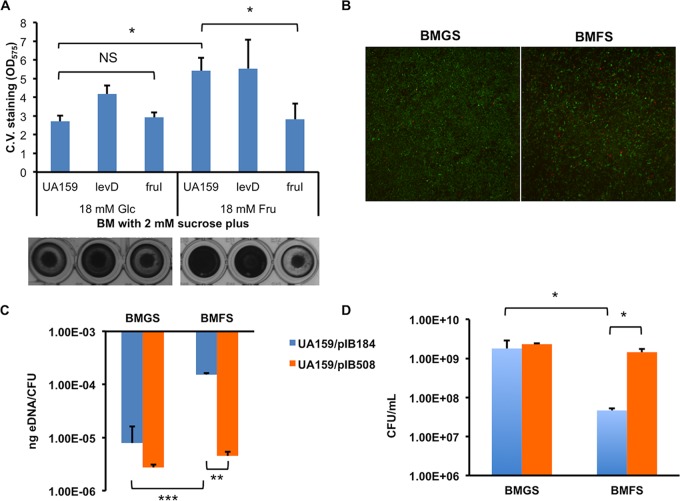
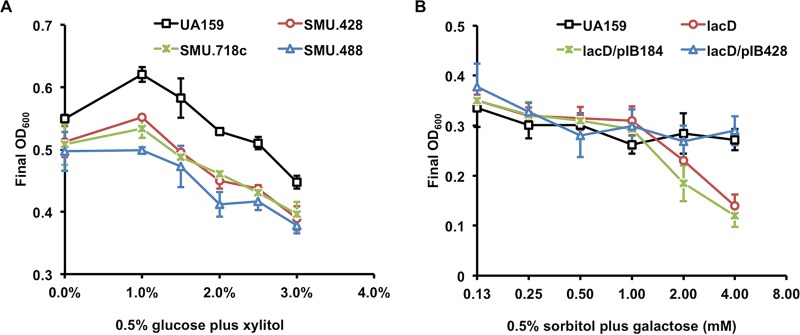
The dental caries pathogen Streptococcus mutans can ferment a variety of sugars to produce organic acids. Exposure of S. mutans to certain nonmetabolizable carbohydrates, such as xylitol, impairs growth and can cause cell death. Recently, the presence of a sugar-phosphate stress in S. mutans was demonstrated using a mutant lacking 1-phosphofructokinase (FruK) that accumulates fructose-1-phosphate (F-1-P). Here, we studied an operon in S. mutans, sppRA, which was highly expressed in the fruK mutant. Biochemical characterization of a recombinant SppA protein indicated that it possessed hexose-phosphate phosphohydrolase activity, with preferences for F-1-P and, to a lesser degree, fructose-6-phosphate (F-6-P). SppA activity was stimulated by Mg2+ and Mn2+ but inhibited by NaF. SppR, a DeoR family regulator, repressed the expression of the sppRA operon to minimum levels in the absence of the fructose-derived metabolite F-1-P and likely also F-6-P. The accumulation of F-1-P, as a result of growth on fructose, not only induced sppA expression, but it significantly altered biofilm maturation through increased cell lysis and enhanced extracellular DNA release. Constitutive expression of sppA, via a plasmid or by deleting sppR, greatly alleviated fructose-induced stress in a fruK mutant, enhanced resistance to xylitol, and reversed the effects of fructose on biofilm formation. Finally, by identifying three additional putative phosphatases that are capable of promoting sugar-phosphate tolerance, we show that S. mutans is capable of mounting a sugar-phosphate stress response by modulating the levels of certain glycolytic intermediates, functions that are interconnected with the ability of the organism to manifest key virulence behaviors.IMPORTANCEStreptococcus mutans is a major etiologic agent for dental caries, primarily due to its ability to form biofilms on the tooth surface and to convert carbohydrates into organic acids. We have discovered a two-gene operon in S. mutans that regulates fructose metabolism by controlling the levels of fructose-1-phosphate, a potential signaling compound that affects bacterial behaviors. With fructose becoming increasingly common and abundant in the human diet, we reveal the ways that fructose may alter bacterial development, stress tolerance, and microbial ecology in the oral cavity to promote oral diseases.
Keywords: biofilm; fructose metabolism; sugar-phosphate phosphohydrolase; sugar-phosphate stress; xylitol.
Copyright © 2018 American Society for Microbiology.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
