

Common Cervicovaginal Microbial Supernatants Alter Cervical Epithelial Function: Mechanisms by Which Lactobacillus crispatus Contributes to Cervical Health
- PMID: 30349508
- PMCID: PMC6186799
- DOI: 10.3389/fmicb.2018.02181
Common Cervicovaginal Microbial Supernatants Alter Cervical Epithelial Function: Mechanisms by Which Lactobacillus crispatus Contributes to Cervical Health
Abstract
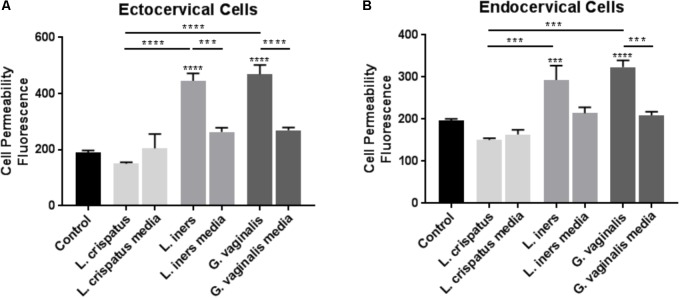
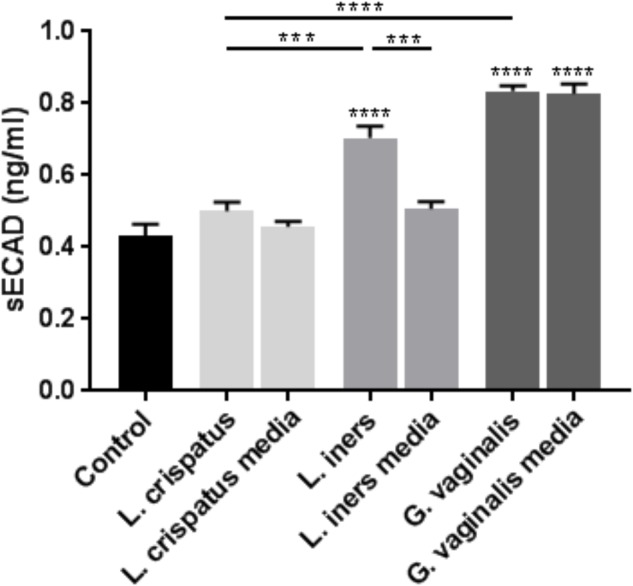
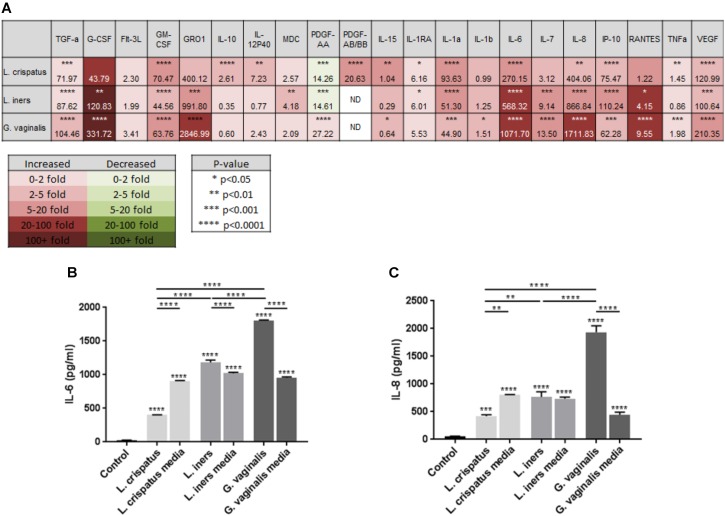
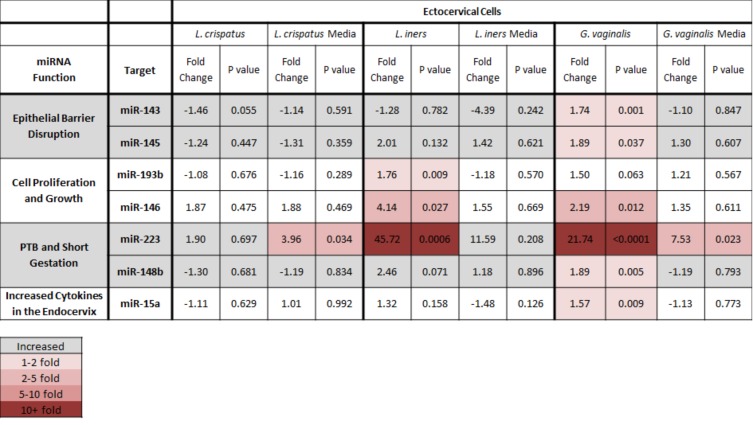
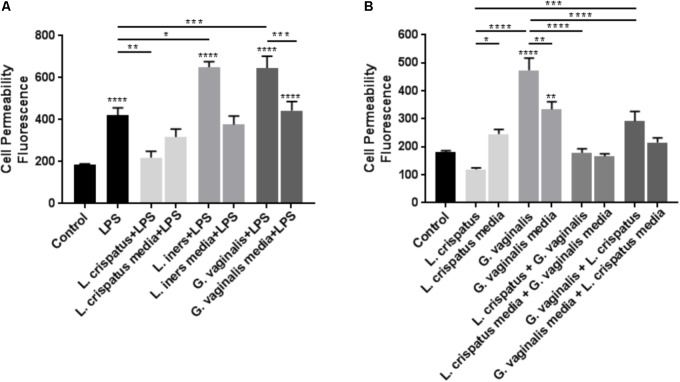
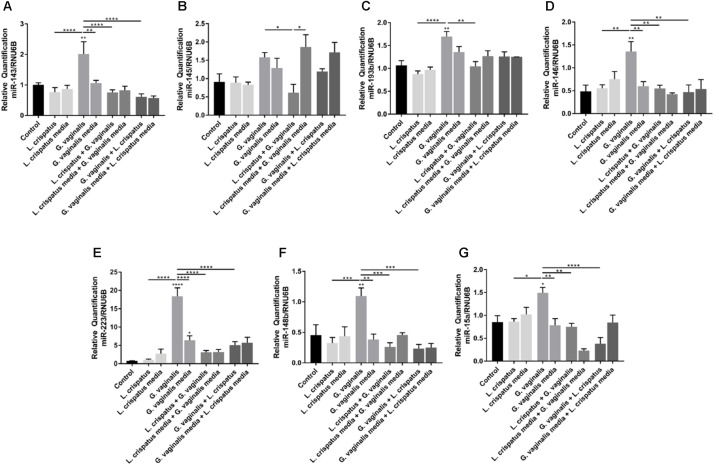
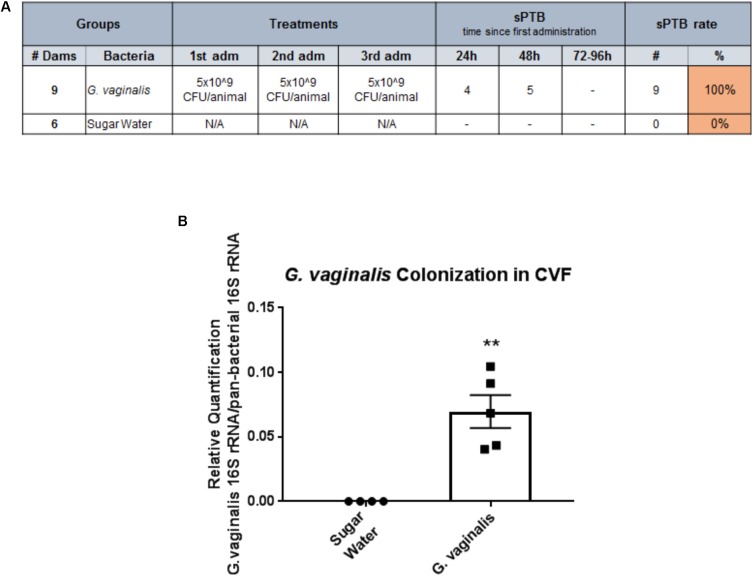
Cervicovaginal (CV) microbiota is associated with vaginal health and disease in non-pregnant women. Recent studies in pregnant women suggest that specific CV microbes are associated with preterm birth (PTB). While the associations between CV microbiota and adverse outcomes have been demonstrated, the mechanisms regulating the associations remain unclear. As the CV space contains an epithelial barrier, we postulate that CV microbiota can alter the epithelial barrier function. We investigated the biological, molecular, and epigenetic effects of Lactobacillus crispatus, Lactobacillus iners, and Gardnerella vaginalis on the cervical epithelial barrier function and determined whether L. crispatus mitigates the effects of lipopolysaccharide (LPS) and G. vaginalis on the cervical epithelial barrier as a possible mechanism by which CV microbiota mitigates disease risk. Ectocervical and endocervical cells treated with L. crispatus, L. iners, and G. vaginalis bacteria-free supernatants alone or combined were used to measure cell permeability, adherens junction proteins, inflammatory mediators, and miRNAs. Ectocervical and endocervical permeability increased after L. iners and G. vaginalis exposure. Soluble epithelial cadherin increased after exposure to L. iners but not G. vaginalis or L. crispatus. A Luminex cytokine/chemokine panel revealed increased proinflammatory mediators in all three bacteria-free supernatants with L. iners and G. vaginalis having more diverse inflammatory effects. L. iners and G. vaginalis altered the expression of cervical-, microbial-, and inflammatory-associated miRNAs. L. crispatus mitigated the LPS or G. vaginalis-induced disruption of the cervical epithelial barrier and reversed the G. vaginalis-mediated increase in miRNA expression. G. vaginalis colonization of the CV space of a pregnant C57/B6 mouse resulted in 100% PTB. These findings demonstrate that L. iners and G. vaginalis alter the cervical epithelial barrier by regulating adherens junction proteins, cervical immune responses, and miRNA expressions. These results provide evidence that L. crispatus confers protection to the cervical epithelial barrier by mitigating LPS- or G. vaginalis-induced miRNAs associated with cervical remodeling, inflammation, and PTB. This study provides further evidence that the CV microbiota plays a role in cervical function by altering the cervical epithelial barrier and initiating PTB. Thus, targeting the CV microbiota and/or its effects on the cervical epithelium may be a potential therapeutic strategy to prevent PTB.
Keywords: Gardnerella vaginalis; Lactobacillus crispatus; Lactobacillus iners; cervix; epithelial barrier; inflammation; miRNA; preterm birth.
Figures
References
-
- Atassi F., Brassart D., Grob P., Graf F., Servin A. L. (2006). Lactobacillus strains isolated from the vaginal microbiota of healthy women inhibit Prevotella bivia and Gardnerella vaginalis in coculture and cell culture. FEMS Immunol. Med. Microbiol. 48 424–432. 10.1111/j.1574-695X.2006.00162.x. - DOI - PubMed
-
- Behrman R., Butler A. (2007). Institute of Medicine (US) Committee on Understanding Premature Birth and Assuring Healthy Outcomes. Preterm Birth: Causes, Consequences, and Prevention. Washington, DC: National Academies Press (US). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases