

Identification of Novel Protein Lysine Acetyltransferases in Escherichia coli
- PMID: 30352934
- PMCID: PMC6199490
- DOI: 10.1128/mBio.01905-18
Identification of Novel Protein Lysine Acetyltransferases in Escherichia coli
Erratum in
-
Correction for Christensen et al., "Identification of Novel Protein Lysine Acetyltransferases in Escherichia coli".mBio. 2019 Apr 9;10(2):e00592-19. doi: 10.1128/mBio.00592-19. mBio. 2019. PMID: 30967468 Free PMC article. No abstract available.
Abstract
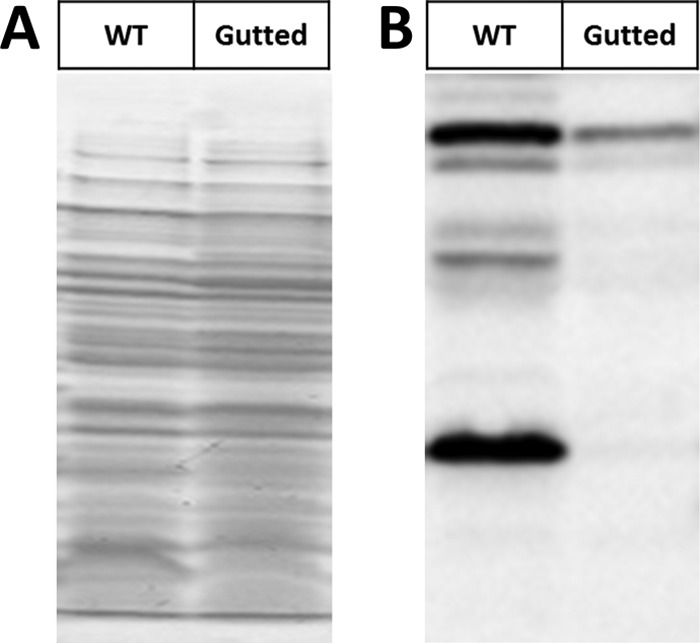
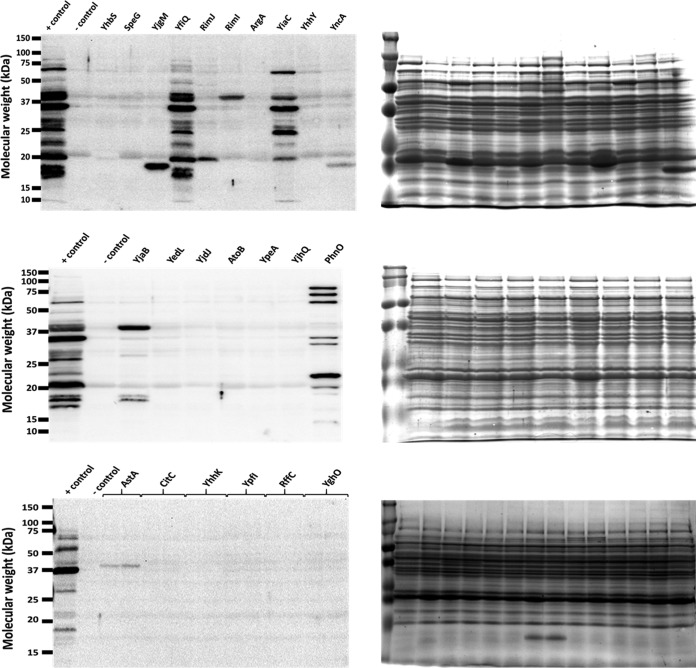
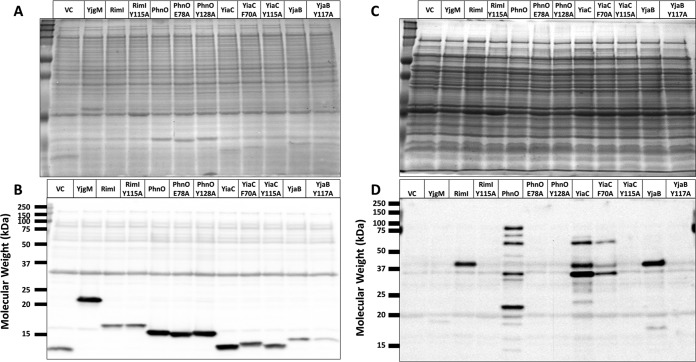
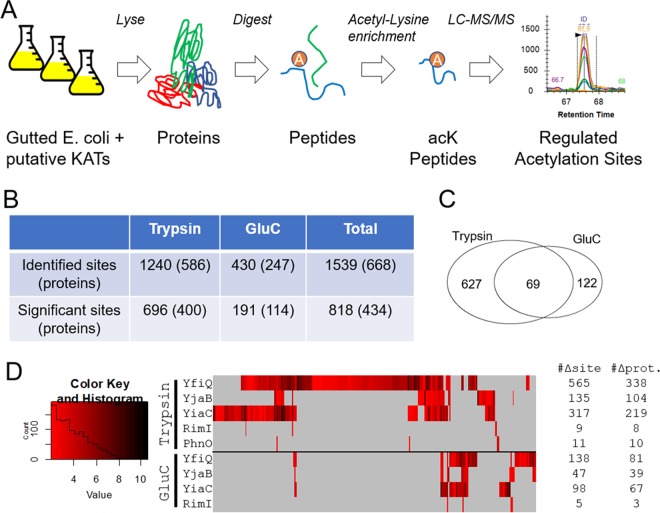
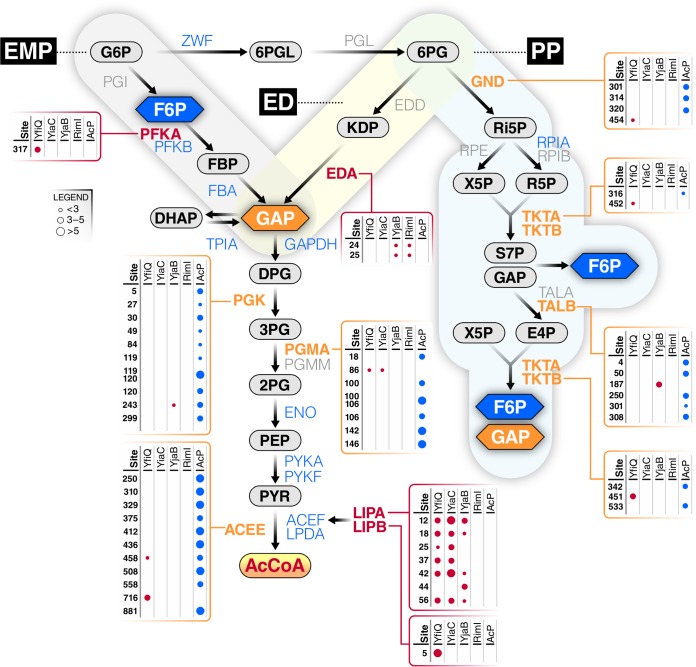
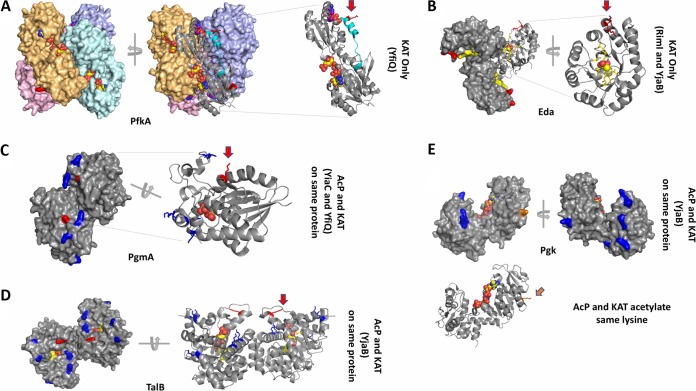
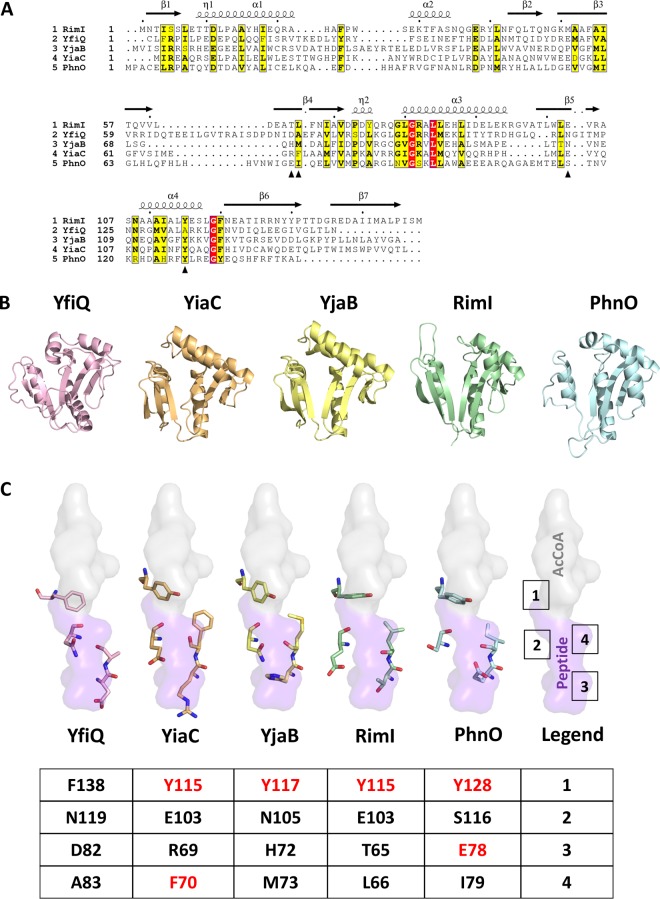
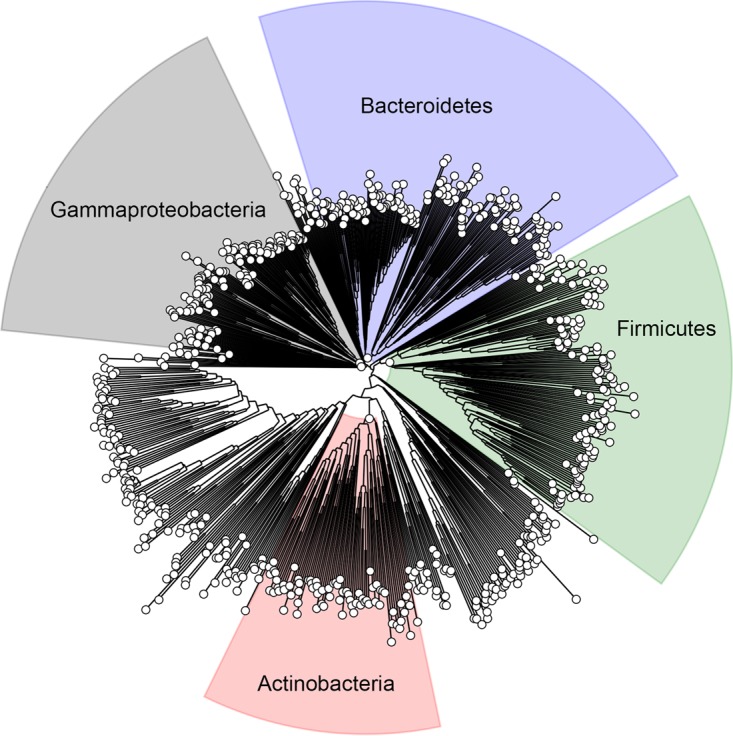
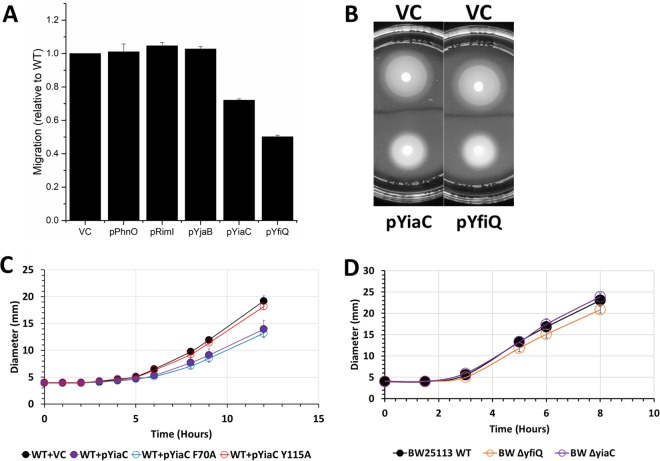
Posttranslational modifications, such as Nε-lysine acetylation, regulate protein function. Nε-lysine acetylation can occur either nonenzymatically or enzymatically. The nonenzymatic mechanism uses acetyl phosphate (AcP) or acetyl coenzyme A (AcCoA) as acetyl donor to modify an Nε-lysine residue of a protein. The enzymatic mechanism uses Nε-lysine acetyltransferases (KATs) to specifically transfer an acetyl group from AcCoA to Nε-lysine residues on proteins. To date, only one KAT (YfiQ, also known as Pka and PatZ) has been identified in Escherichia coli Here, we demonstrate the existence of 4 additional E. coli KATs: RimI, YiaC, YjaB, and PhnO. In a genetic background devoid of all known acetylation mechanisms (most notably AcP and YfiQ) and one deacetylase (CobB), overexpression of these putative KATs elicited unique patterns of protein acetylation. We mutated key active site residues and found that most of them eliminated enzymatic acetylation activity. We used mass spectrometry to identify and quantify the specificity of YfiQ and the four novel KATs. Surprisingly, our analysis revealed a high degree of substrate specificity. The overlap between KAT-dependent and AcP-dependent acetylation was extremely limited, supporting the hypothesis that these two acetylation mechanisms play distinct roles in the posttranslational modification of bacterial proteins. We further showed that these novel KATs are conserved across broad swaths of bacterial phylogeny. Finally, we determined that one of the novel KATs (YiaC) and the known KAT (YfiQ) can negatively regulate bacterial migration. Together, these results emphasize distinct and specific nonenzymatic and enzymatic protein acetylation mechanisms present in bacteria.IMPORTANCENε-Lysine acetylation is one of the most abundant and important posttranslational modifications across all domains of life. One of the best-studied effects of acetylation occurs in eukaryotes, where acetylation of histone tails activates gene transcription. Although bacteria do not have true histones, Nε-lysine acetylation is prevalent; however, the role of these modifications is mostly unknown. We constructed an E. coli strain that lacked both known acetylation mechanisms to identify four new Nε-lysine acetyltransferases (RimI, YiaC, YjaB, and PhnO). We used mass spectrometry to determine the substrate specificity of these acetyltransferases. Structural analysis of selected substrate proteins revealed site-specific preferences for enzymatic acetylation that had little overlap with the preferences of the previously reported acetyl-phosphate nonenzymatic acetylation mechanism. Finally, YiaC and YfiQ appear to regulate flagellum-based motility, a phenotype critical for pathogenesis of many organisms. These acetyltransferases are highly conserved and reveal deeper and more complex roles for bacterial posttranslational modification.
Keywords: GNAT; acetylation; acetyltransferase; bacteria; lysine acetyltransferase; mass spectrometry; protein acetyltransferase; proteomics.
Copyright © 2018 Christensen et al.
Figures
References
-
- Kuhn ML, Zemaitaitis B, Hu LI, Sahu A, Sorensen D, Minasov G, Lima BP, Scholle M, Mrksich M, Anderson WF, Gibson BW, Schilling B, Wolfe AJ. 2014. Structural, kinetic and proteomic characterization of acetyl phosphate-dependent bacterial protein acetylation. PLoS One 9:e94816. doi: 10.1371/journal.pone.0094816. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
