

miR-146a Suppresses SUMO1 Expression and Induces Cardiac Dysfunction in Maladaptive Hypertrophy
- PMID: 30355233
- PMCID: PMC6205728
- DOI: 10.1161/CIRCRESAHA.118.312751
miR-146a Suppresses SUMO1 Expression and Induces Cardiac Dysfunction in Maladaptive Hypertrophy
Abstract
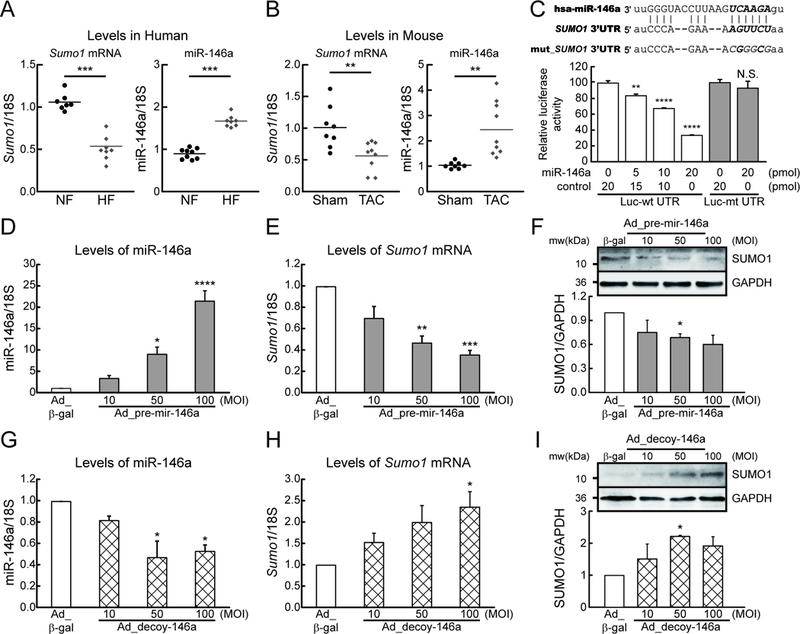
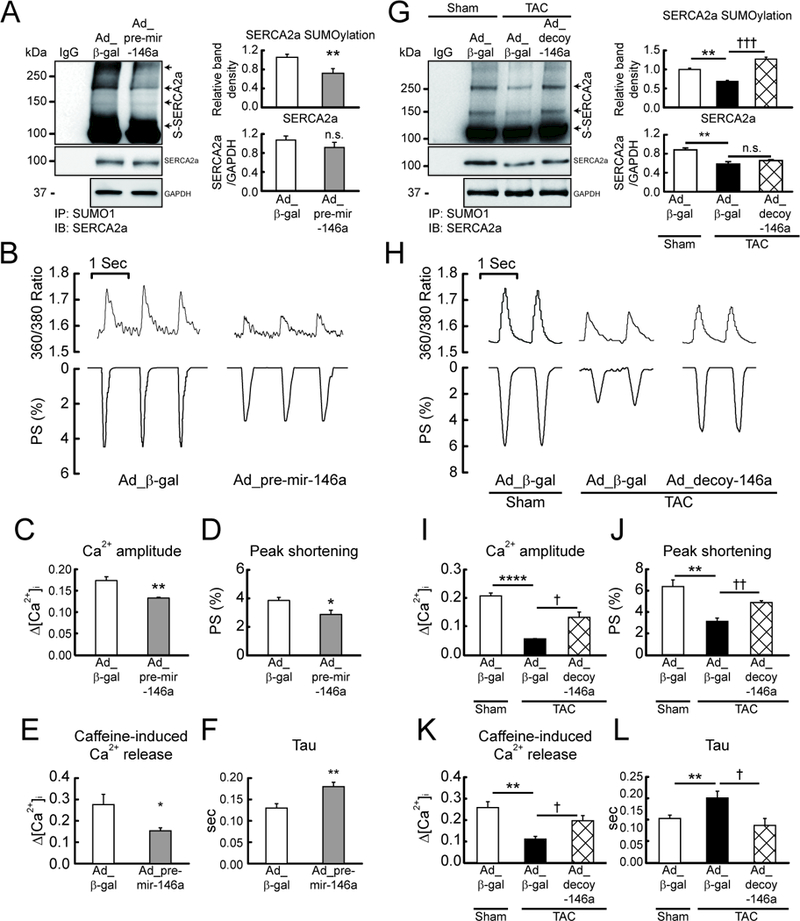
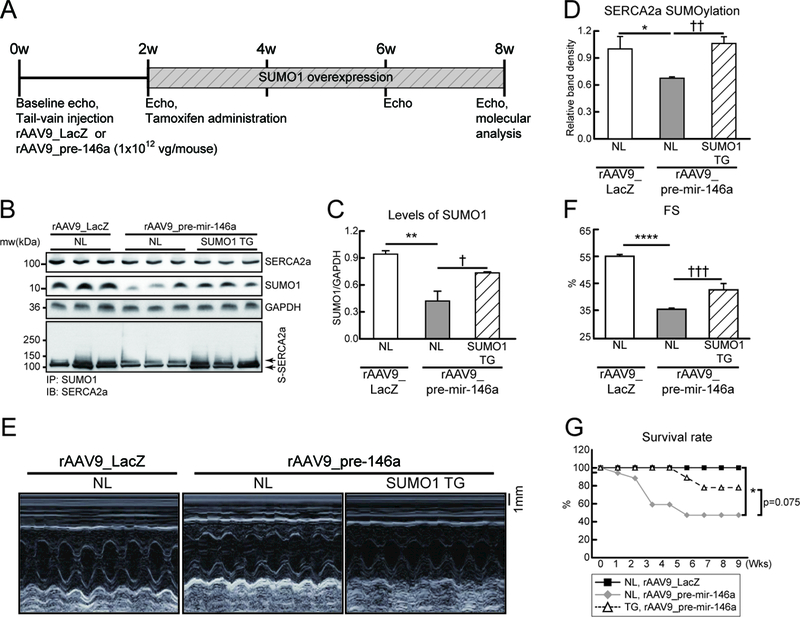
Rationale: Abnormal SUMOylation has emerged as a characteristic of heart failure (HF) pathology. Previously, we found reduced SUMO1 (small ubiquitin-like modifier 1) expression and SERCA2a (sarcoplasmic reticulum Ca2+-ATPase) SUMOylation in human and animal HF models. SUMO1 gene delivery or small molecule activation of SUMOylation restored SERCA2a SUMOylation and cardiac function in HF models. Despite the critical role of SUMO1 in HF, the regulatory mechanisms underlying SUMO1 expression are largely unknown.
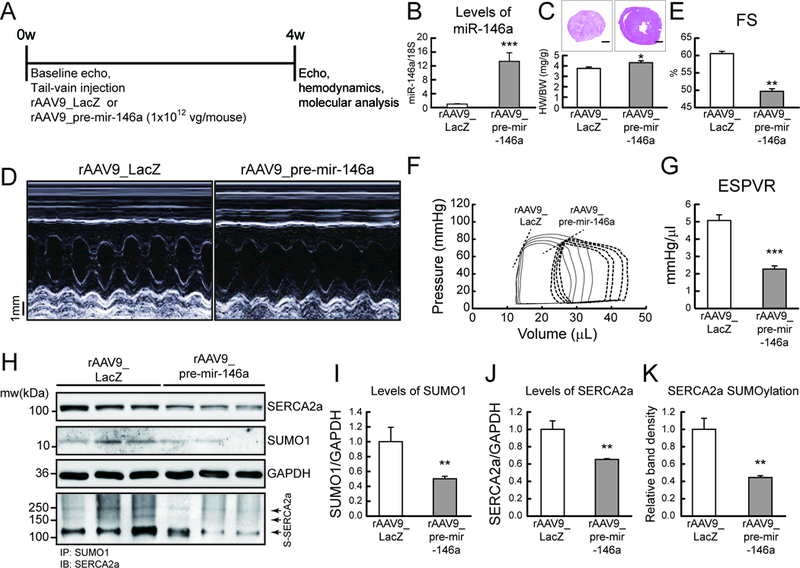
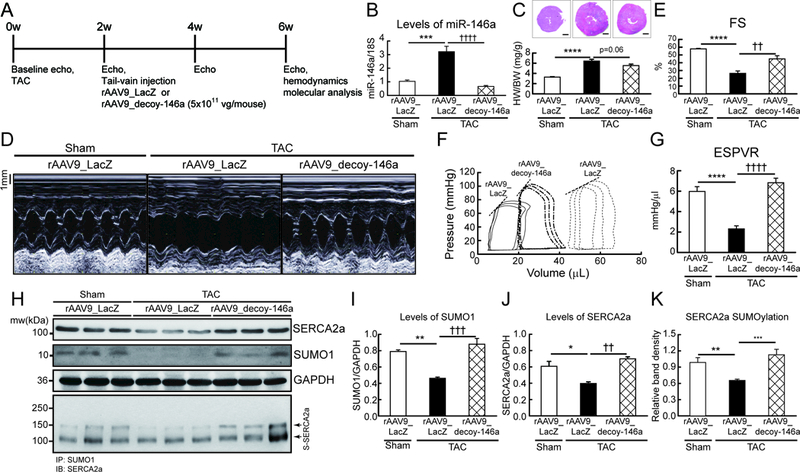
Objective: To examine miR-146a-mediated SUMO1 regulation and its consequent effects on cardiac morphology and function.
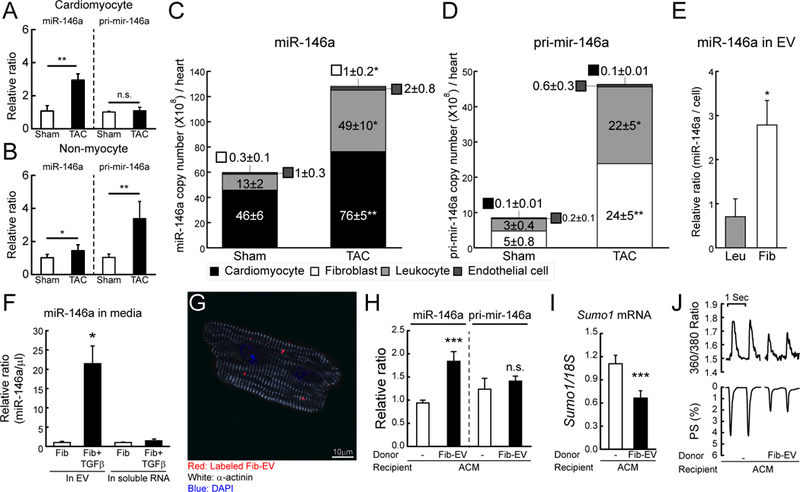
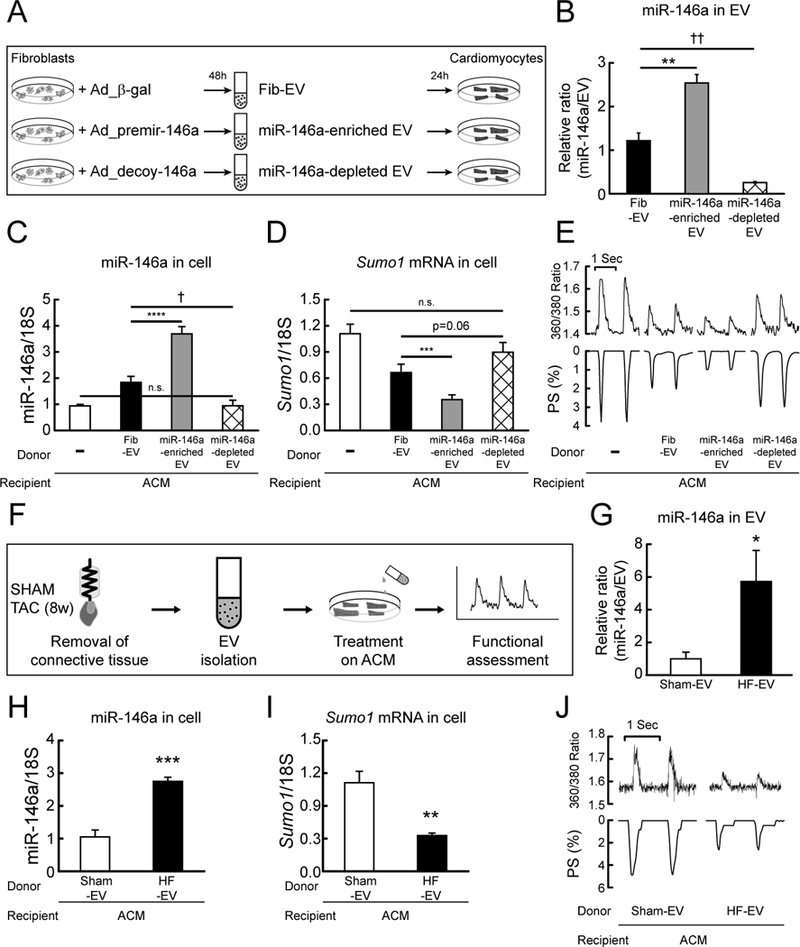
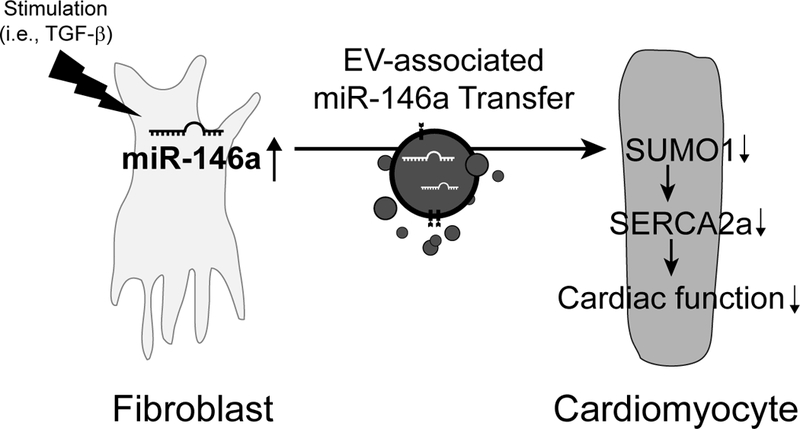
Methods and results: In this study, miR-146a was identified as a SUMO1-targeting microRNA in the heart. A strong correlation was observed between miR-146a and SUMO1 expression in failing mouse and human hearts. miR-146a was manipulated in cardiomyocytes through AAV9 (adeno-associated virus serotype 9)-mediated gene delivery, and cardiac morphology and function were analyzed by echocardiography and hemodynamics. Overexpression of miR-146a reduced SUMO1 expression, SERCA2a SUMOylation, and cardiac contractility in vitro and in vivo. The effects of miR-146a inhibition on HF pathophysiology were examined by transducing a tough decoy of miR-146a into mice subjected to transverse aortic constriction. miR-146a inhibition improved cardiac contractile function and normalized SUMO1 expression. The regulatory mechanisms of miR-146a upregulation were elucidated by examining the major miR-146a-producing cell types and transfer mechanisms. Notably, transdifferentiation of fibroblasts triggered miR-146a overexpression and secretion through extracellular vesicles, and the extracellular vesicle-associated miR-146a transfer was identified as the causative mechanism of miR-146a upregulation in failing cardiomyocytes. Finally, extracellular vesicles isolated from failing hearts were shown to contain high levels of miR-146a and exerted negative effects on the SUMO1/SERCA2a signaling axis and hence cardiomyocyte contractility.
Conclusions: Taken together, our results show that miR-146a is a novel regulator of the SUMOylation machinery in the heart, which can be targeted for therapeutic intervention.
Keywords: extracellular vesicle; heart failure; mice; microRNA; sarcoplasmic reticulum.
Figures
References
-
- Mozaffarian D, Benjamin EJ, Go AS, Arnett DK, Blaha MJ, Cushman M, Das SR, de Ferranti S, Despres JP, Fullerton HJ, Howard VJ, Huffman MD, Isasi CR, Jimenez MC, Judd SE, Kissela BM, Lichtman JH, Lisabeth LD, Liu S, Mackey RH, Magid DJ, McGuire DK, Mohler ER, 3rd, Moy CS, Muntner P, Mussolino ME, Nasir K, Neumar RW, Nichol G, Palaniappan L, Pandey DK, Reeves MJ, Rodriguez CJ, Rosamond W, Sorlie PD, Stein J, Towfighi A, Turan TN, Virani SS, Woo D, Yeh RW, Turner MB. Heart Disease and Stroke Statistics-2016 Update: A Report From the American Heart Association. Circulation. 2016;133(4):e38–360. - PubMed
-
- Miyamoto MI, del Monte F, Schmidt U, DiSalvo TS, Kang ZB, Matsui T, Guerrero JL, Gwathmey JK, Rosenzweig A, Hajjar RJ. Adenoviral gene transfer of SERCA2a improves left-ventricular function in aortic-banded rats in transition to heart failure. Proc Natl Acad Sci U S A. 2000;97(2):793–798. - PMC - PubMed
-
- del Monte F, Hajjar RJ, Harding SE. Overwhelming evidence of the beneficial effects of SERCA gene transfer in heart failure. Circ Res. 2001;88(11):E66–67. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL131404/HL/NHLBI NIH HHS/United States
- P50 HL112324/HL/NHLBI NIH HHS/United States
- R01 HL119046/HL/NHLBI NIH HHS/United States
- R01 HL128099/HL/NHLBI NIH HHS/United States
- R01 HL117505/HL/NHLBI NIH HHS/United States
- R00 HL116645/HL/NHLBI NIH HHS/United States
- T32 HL007824/HL/NHLBI NIH HHS/United States
- R01 HL135093/HL/NHLBI NIH HHS/United States
- 17POST33410877/AHA/American Heart Association-American Stroke Association/United States
- K99 HL116645/HL/NHLBI NIH HHS/United States
- R01 HL129814/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
