

Multiple tumor suppressors regulate a HIF-dependent negative feedback loop via ISGF3 in human clear cell renal cancer
- PMID: 30355451
- PMCID: PMC6234029
- DOI: 10.7554/eLife.37925
Multiple tumor suppressors regulate a HIF-dependent negative feedback loop via ISGF3 in human clear cell renal cancer
Erratum in
-
Correction: Multiple tumor suppressors regulate a HIF-dependent negative feedback loop via ISGF3 in human clear cell renal cancer.Elife. 2021 Apr 21;10:e69256. doi: 10.7554/eLife.69256. Elife. 2021. PMID: 33881395 Free PMC article.
Abstract
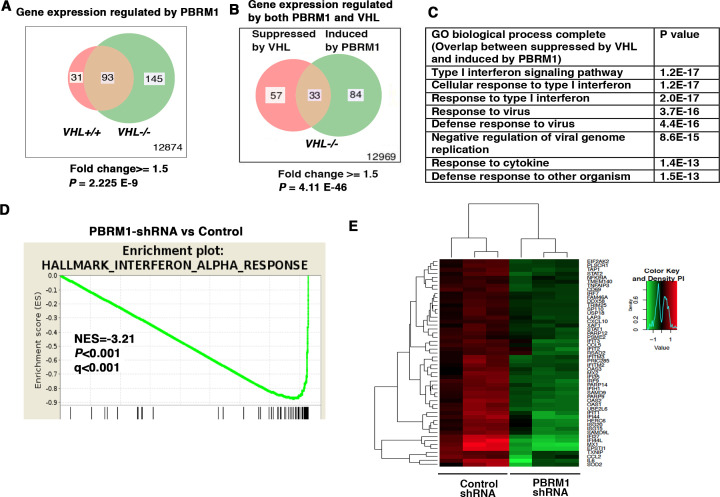
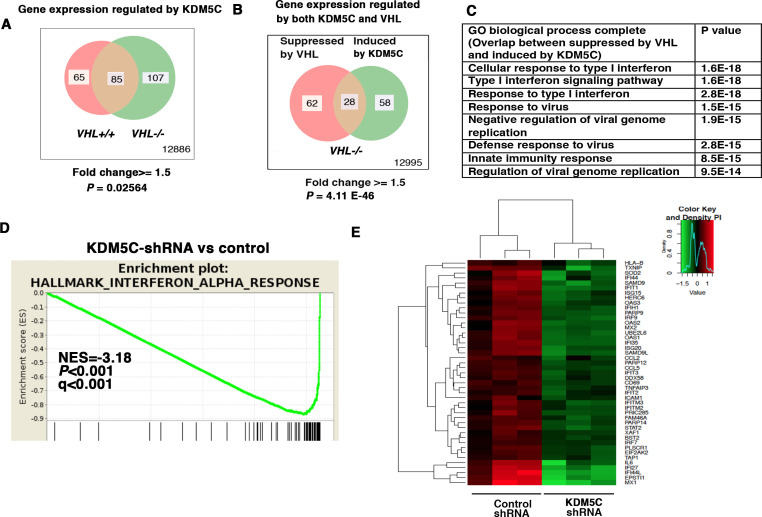
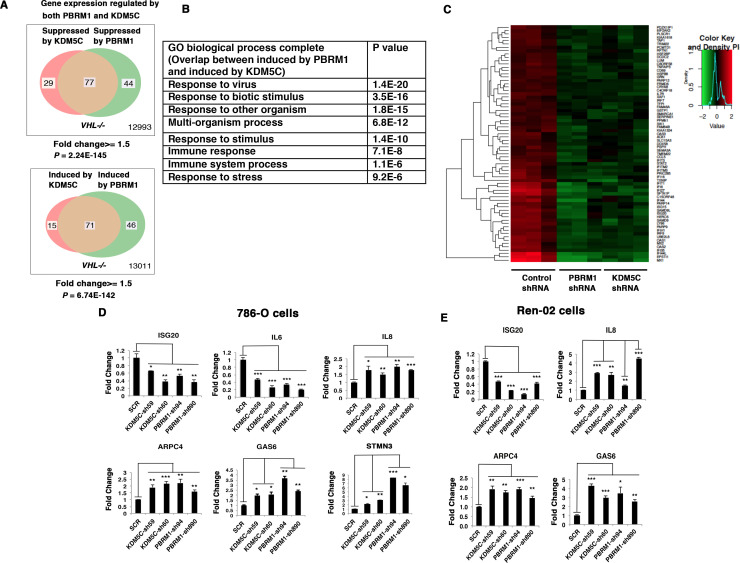
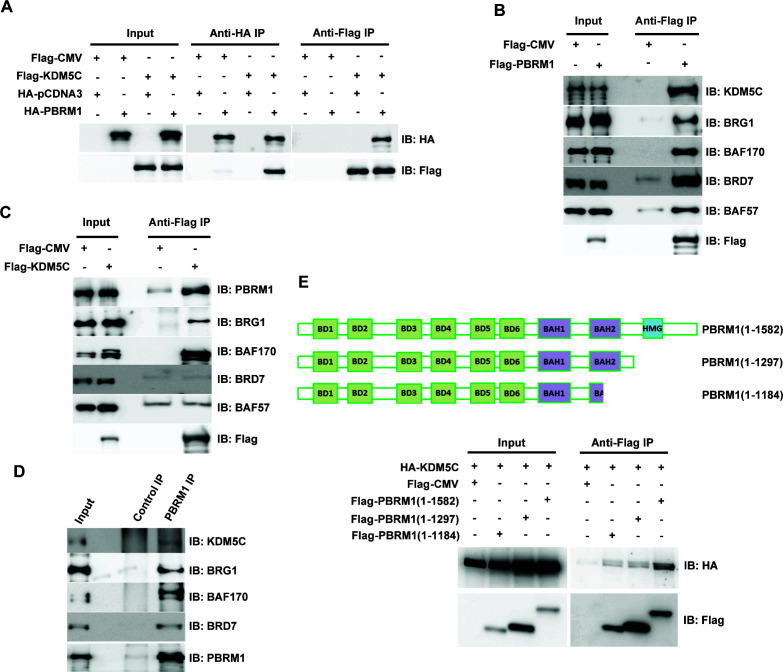
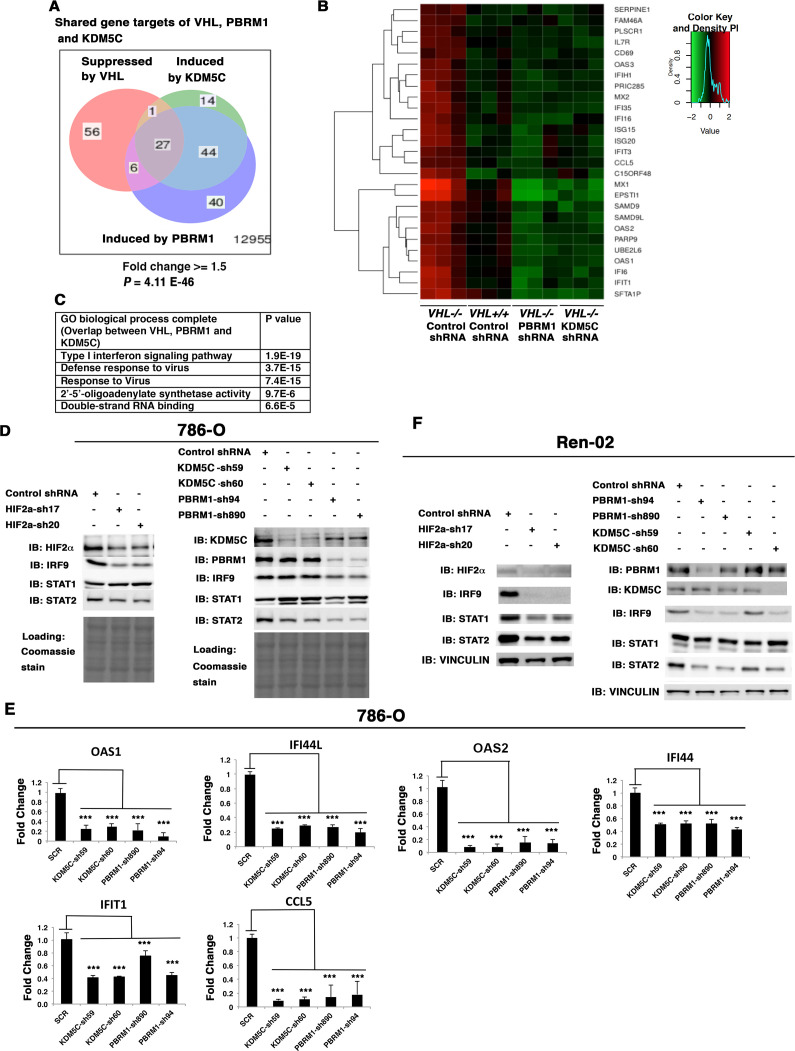
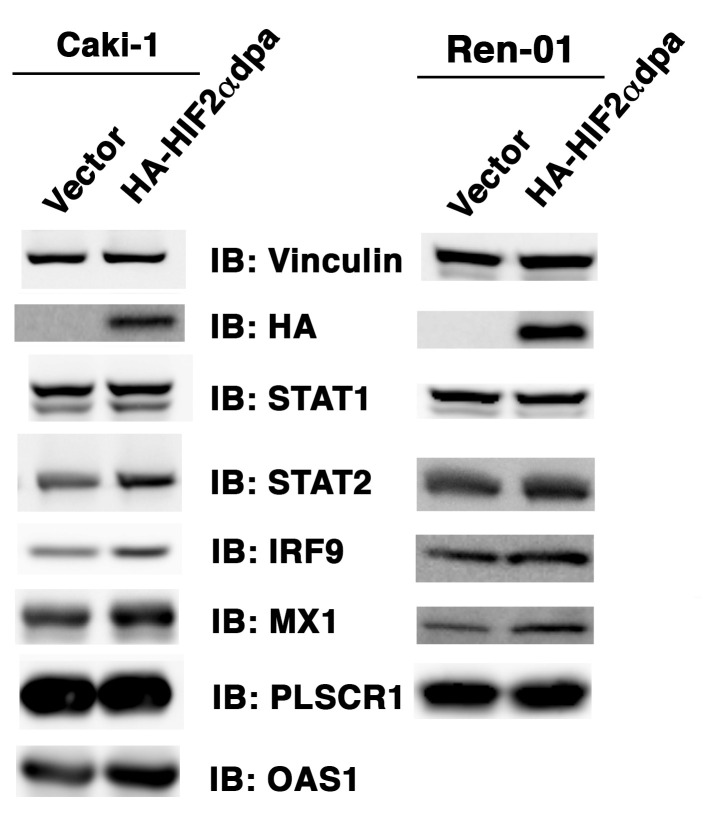
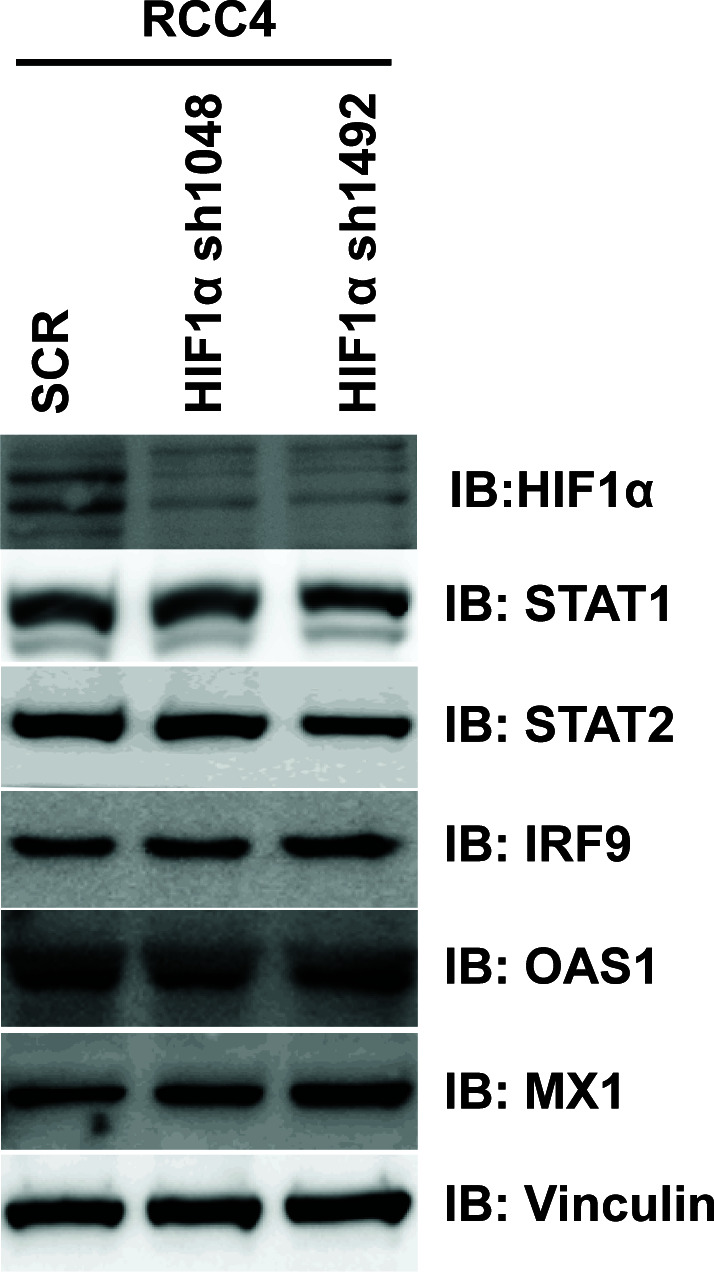
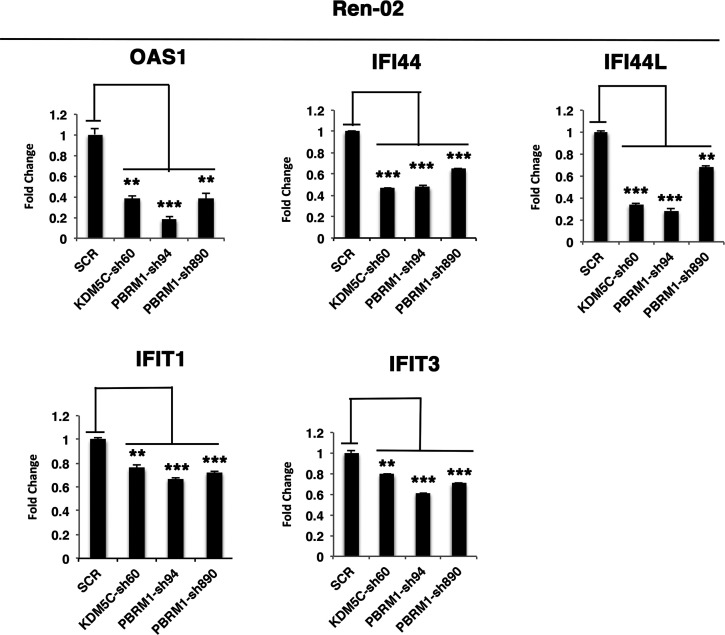
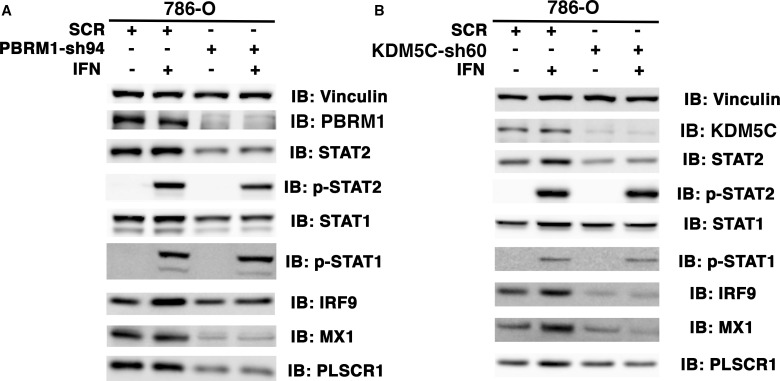
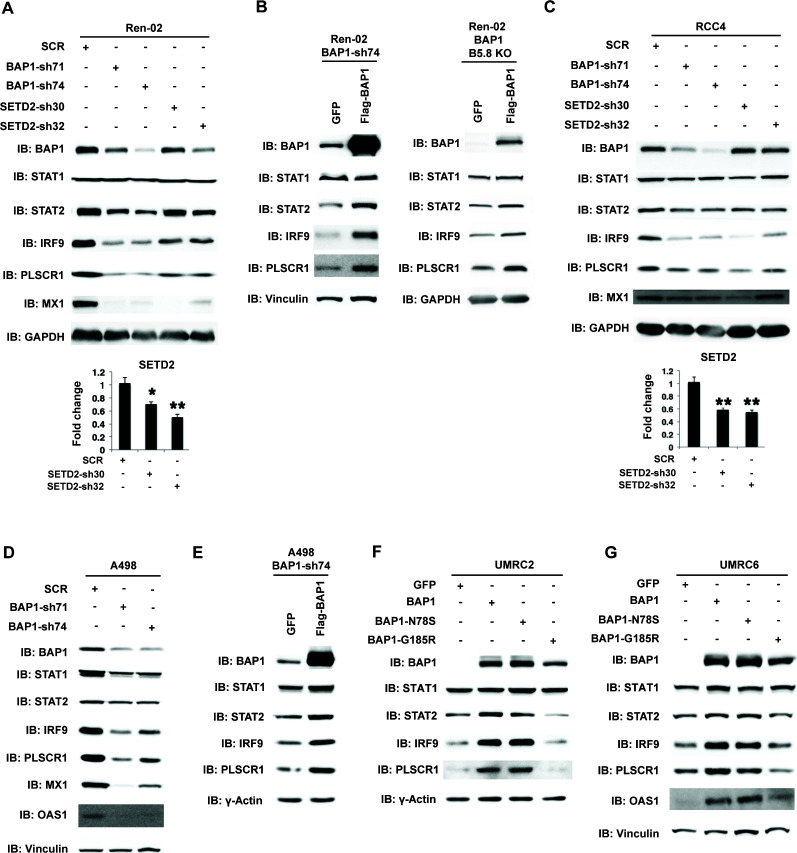
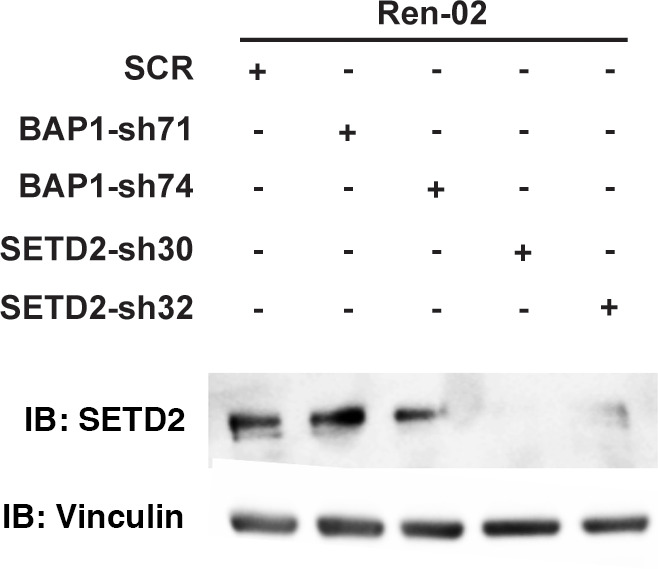
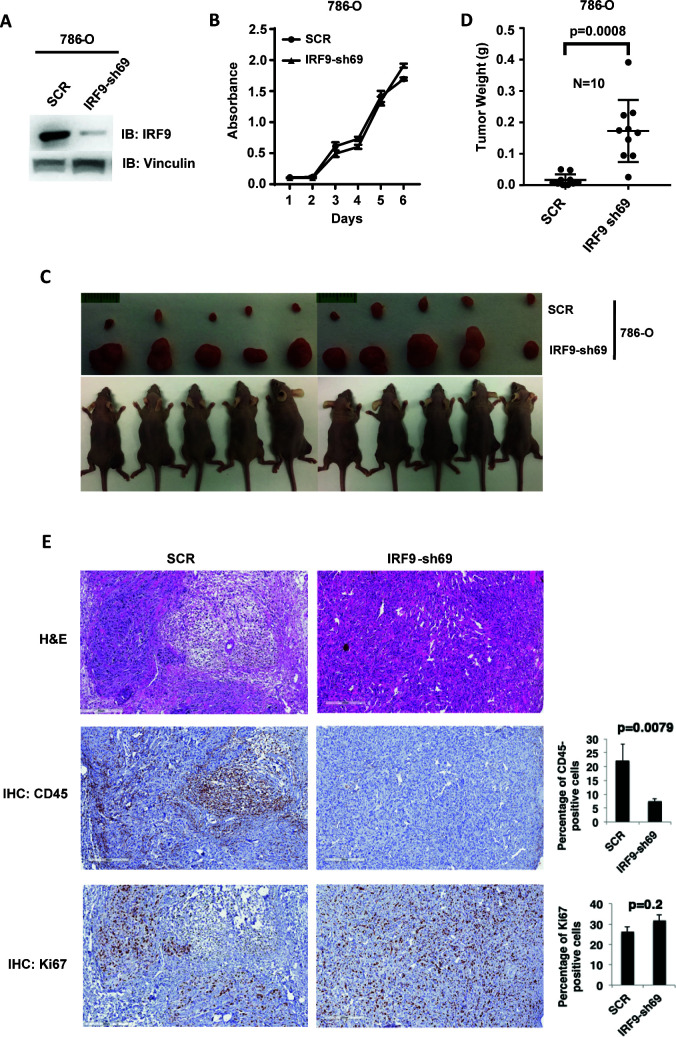
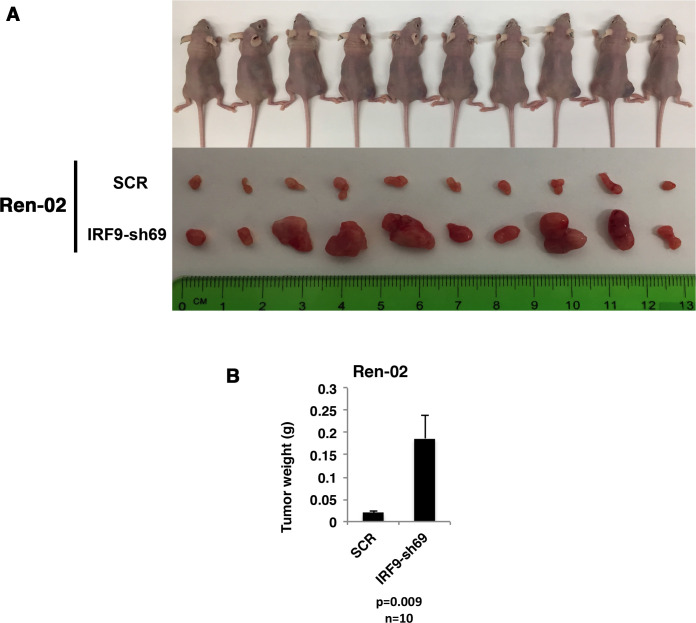
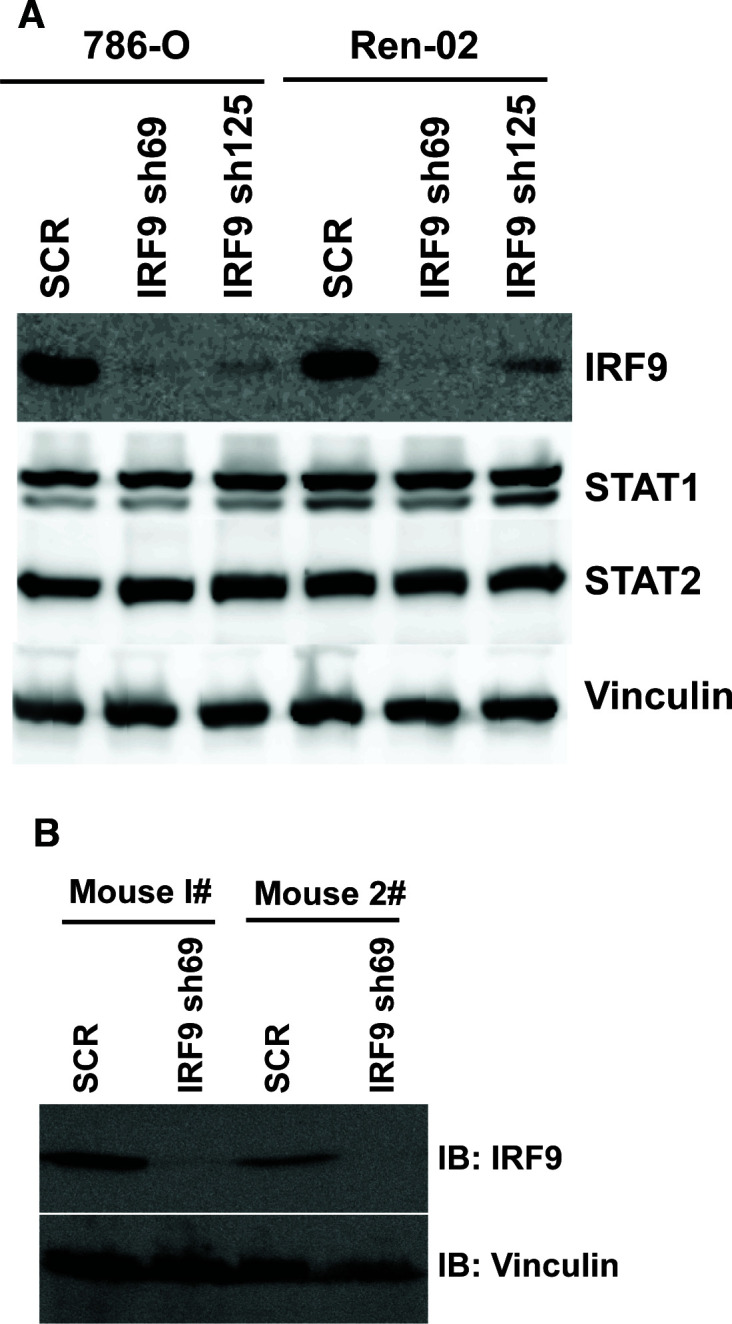
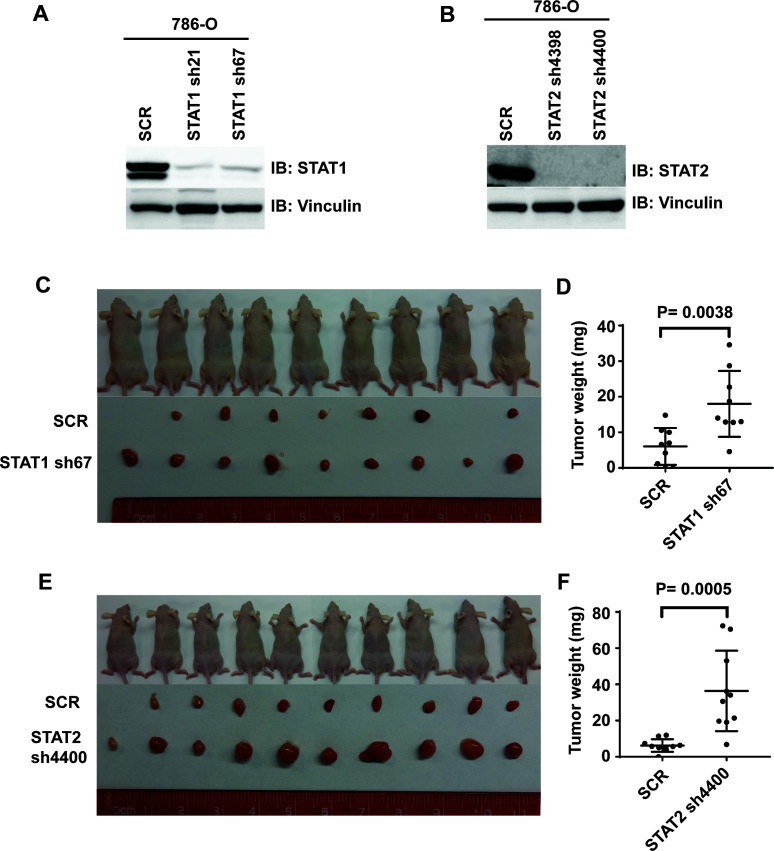
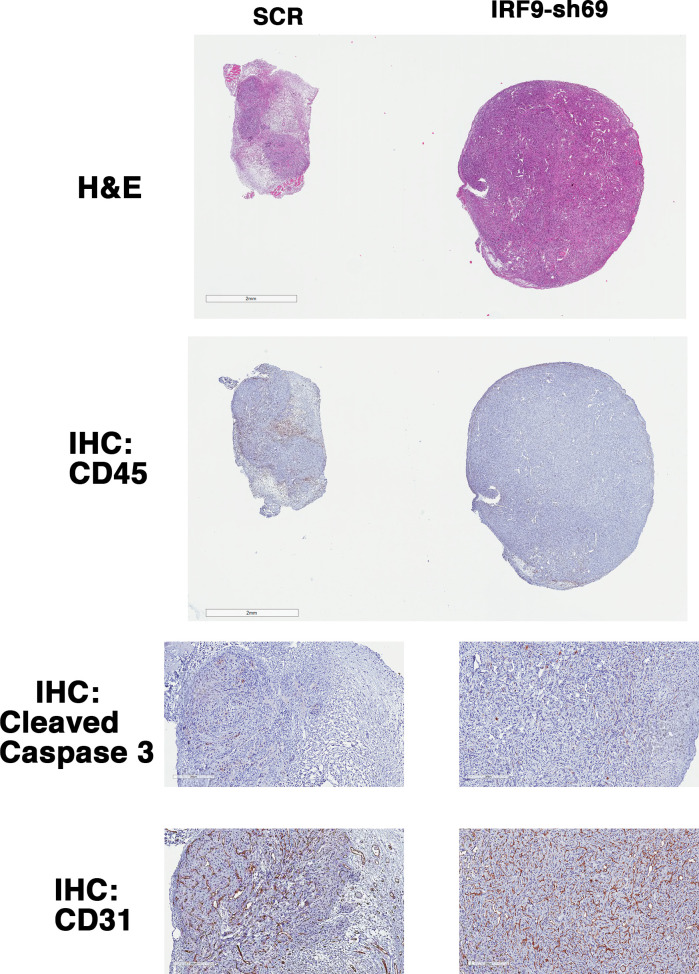
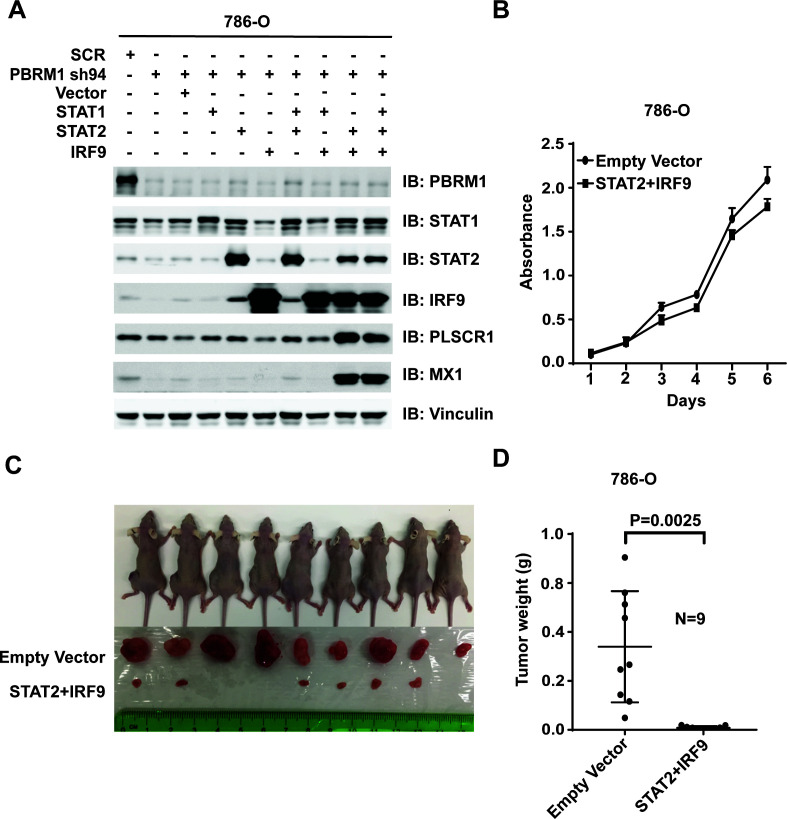
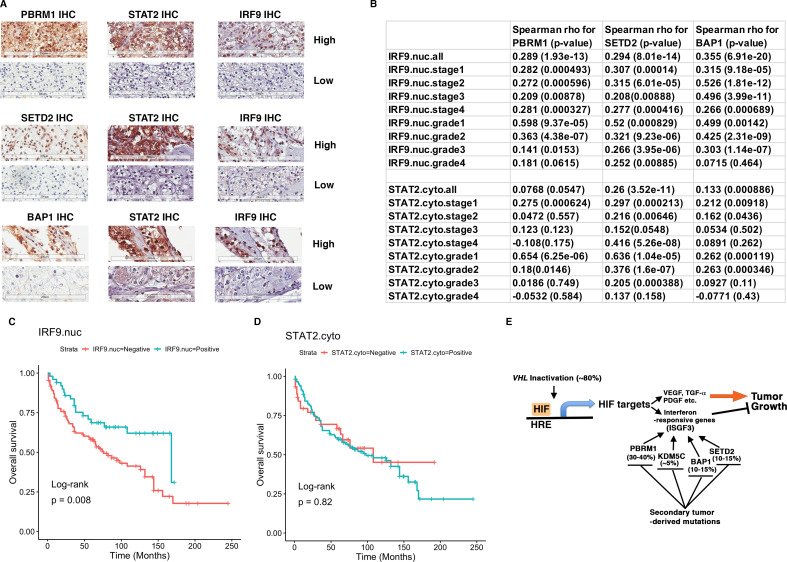
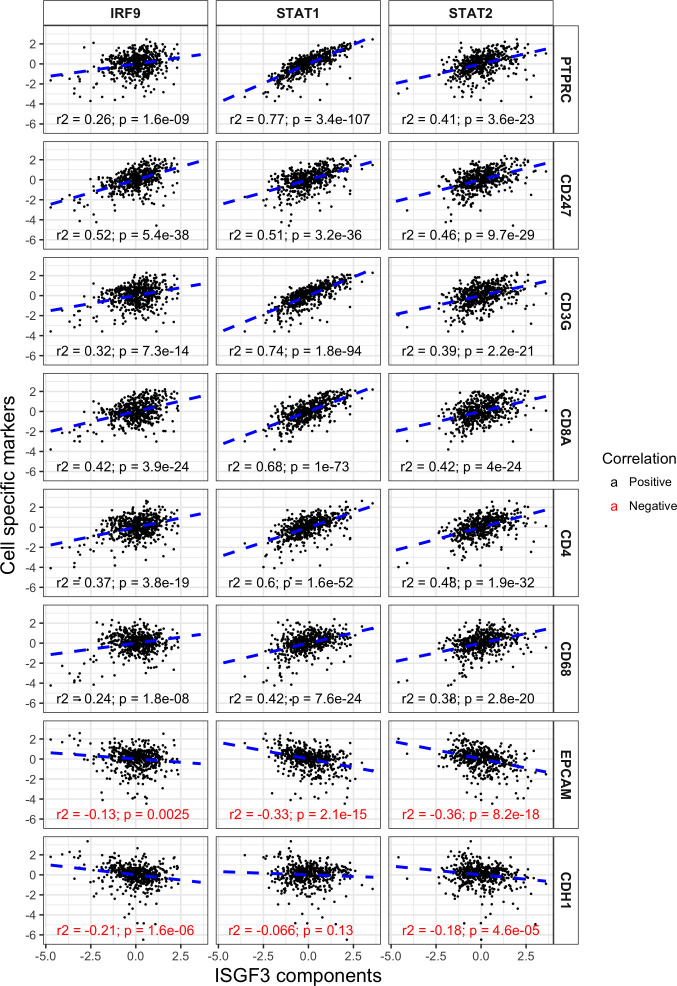
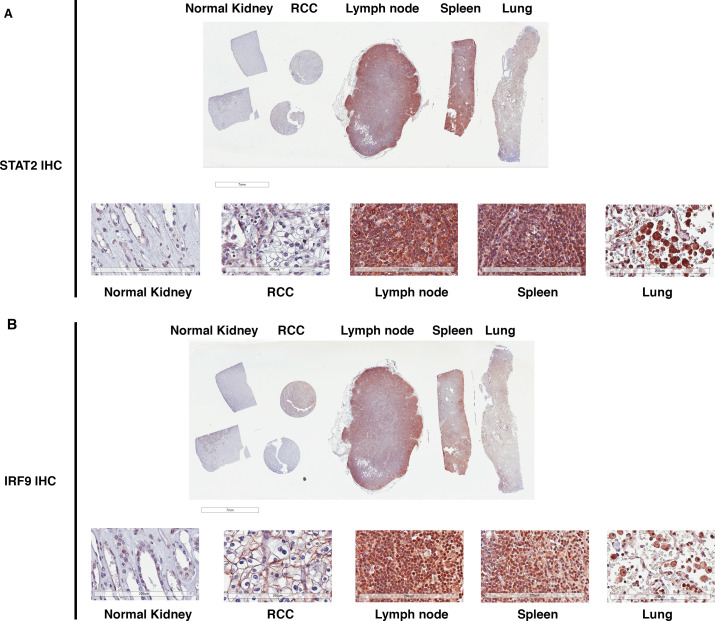
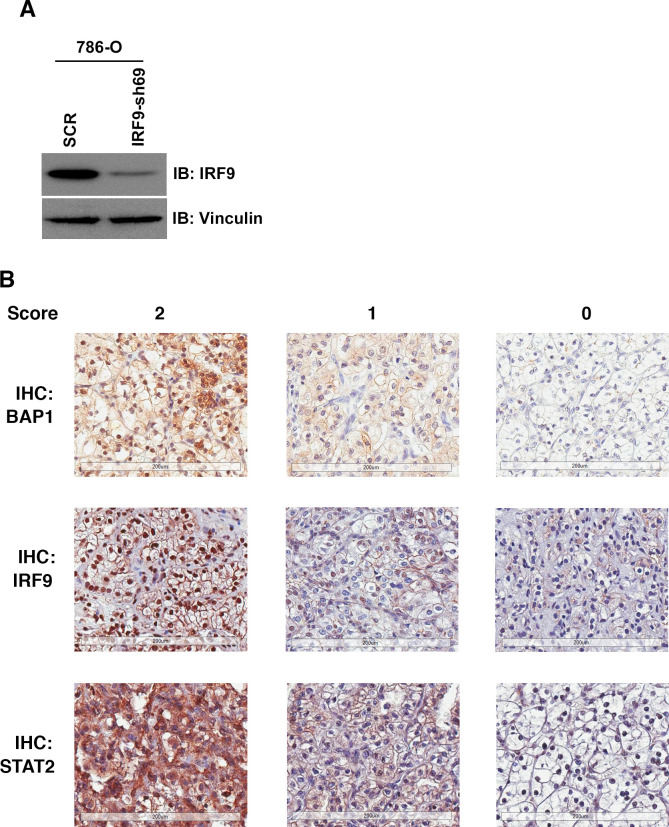
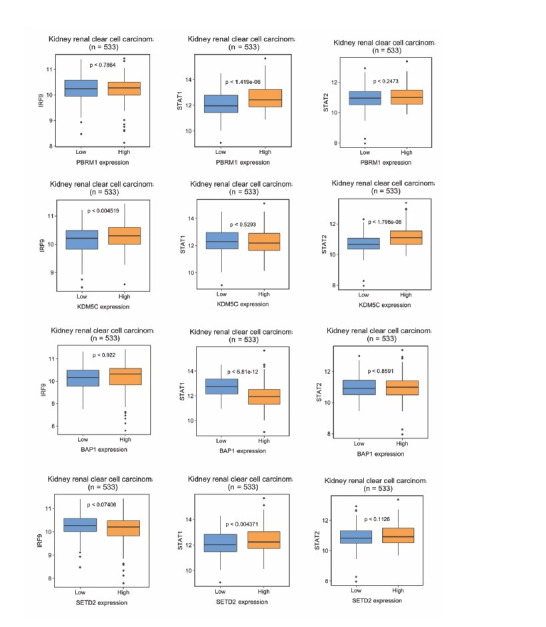
Whereas VHL inactivation is a primary event in clear cell renal cell carcinoma (ccRCC), the precise mechanism(s) of how this interacts with the secondary mutations in tumor suppressor genes, including PBRM1, KDM5C/JARID1C, SETD2, and/or BAP1, remains unclear. Gene expression analyses reveal that VHL, PBRM1, or KDM5C share a common regulation of interferon response expression signature. Loss of HIF2α, PBRM1, or KDM5C in VHL-/-cells reduces the expression of interferon stimulated gene factor 3 (ISGF3), a transcription factor that regulates the interferon signature. Moreover, loss of SETD2 or BAP1 also reduces the ISGF3 level. Finally, ISGF3 is strongly tumor-suppressive in a xenograft model as its loss significantly enhances tumor growth. Conversely, reactivation of ISGF3 retards tumor growth by PBRM1-deficient ccRCC cells. Thus after VHL inactivation, HIF induces ISGF3, which is reversed by the loss of secondary tumor suppressors, suggesting that this is a key negative feedback loop in ccRCC.
Keywords: BAP1; ISGF3; KDM5C; PBRM1; VHL; cancer biology; chromosomes; gene expression; human; kidney cancer.
© 2018, Liao et al.
Conflict of interest statement
LL, ZL, LL, WC, EC, JN, XN, WJ, ZZ, WC, GJ, ED, JT, RU, YW, GS, JS, SP, YX, QY, HY No competing interests declared
Figures
Comment in
-
A central anti-oncogenic pathway in ccRCC.Nat Rev Nephrol. 2019 Jan;15(1):2. doi: 10.1038/s41581-018-0083-6. Nat Rev Nephrol. 2019. PMID: 30429588 No abstract available.
References
-
- Abdel-Rahman MH, Pilarski R, Cebulla CM, Massengill JB, Christopher BN, Boru G, Hovland P, Davidorf FH. Germline BAP1 mutation predisposes to Uveal Melanoma, lung adenocarcinoma, Meningioma, and other cancers. Journal of Medical Genetics. 2011;48:856–859. doi: 10.1136/jmedgenet-2011-100156. - DOI - PMC - PubMed
-
- Abernathy A. U.S. Food and Drug Administration (FDA) Accepts Bristol-Myers Squibb’s Application for Opdivo (nivolumab) Plus Yervoy (ipilimumab) in Intermediate- and Poor-Risk Patients with Advanced Renal Cell Carcinoma and Grants Priority Review. [December 13, 2017];2017 https://investors.bms.com/iframes/press-releases/press-release-details/2...
-
- Benci JL, Xu B, Qiu Y, Wu TJ, Dada H, Twyman-Saint Victor C, Cucolo L, Lee DSM, Pauken KE, Huang AC, Gangadhar TC, Amaravadi RK, Schuchter LM, Feldman MD, Ishwaran H, Vonderheide RH, Maity A, Wherry EJ, Minn AJ. Tumor interferon signaling regulates a multigenic resistance program to immune checkpoint blockade. Cell. 2016;167:1540–1554. doi: 10.1016/j.cell.2016.11.022. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
