

CRISPR/Cas9-mediated homology-directed repair by ssODNs in zebrafish induces complex mutational patterns resulting from genomic integration of repair-template fragments
- PMID: 30355591
- PMCID: PMC6215429
- DOI: 10.1242/dmm.035352
CRISPR/Cas9-mediated homology-directed repair by ssODNs in zebrafish induces complex mutational patterns resulting from genomic integration of repair-template fragments
Abstract
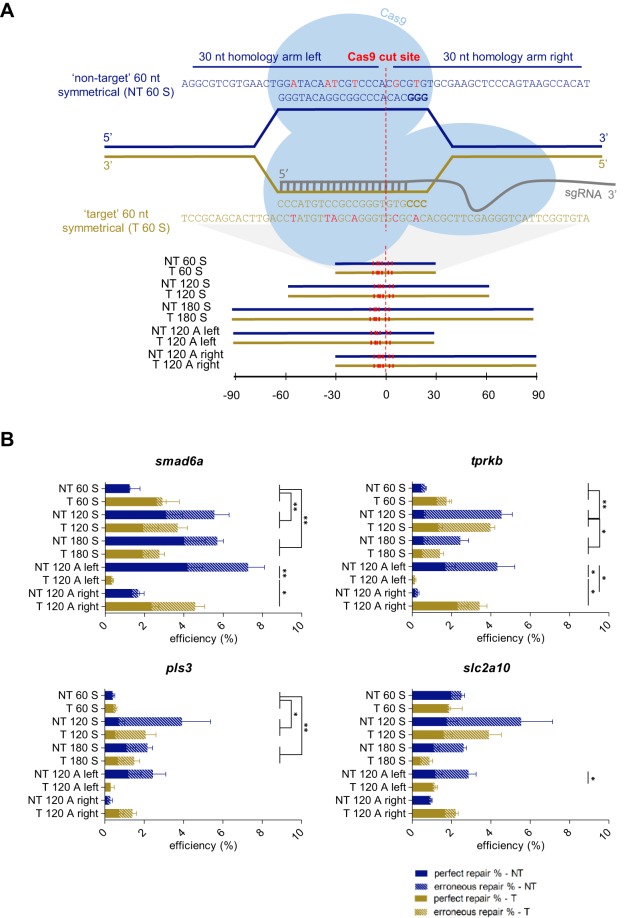
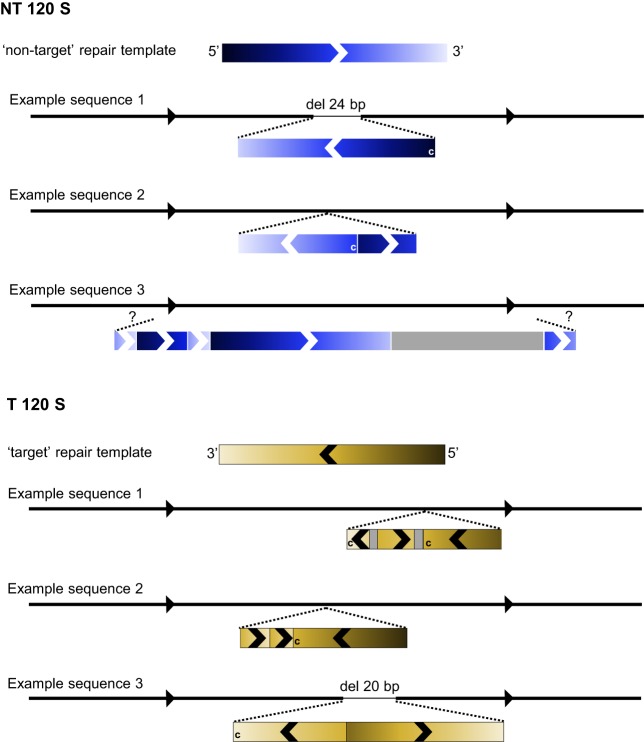
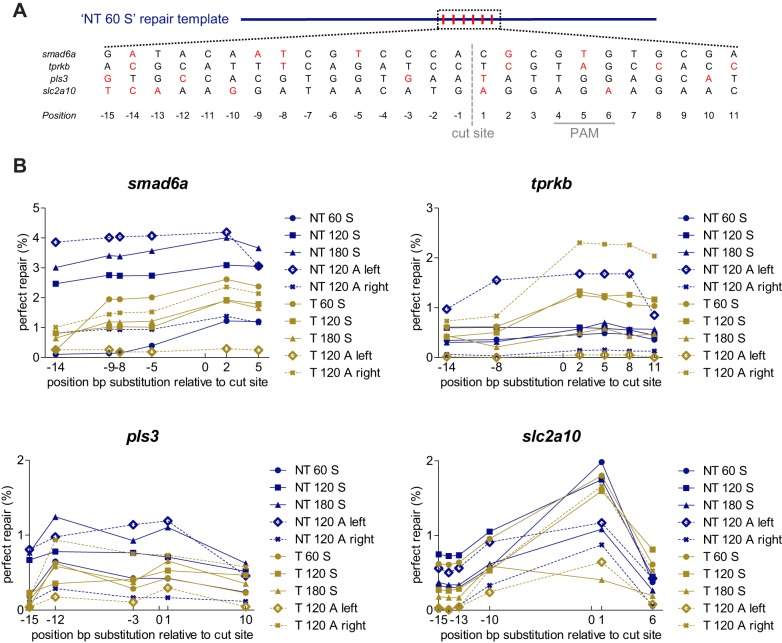
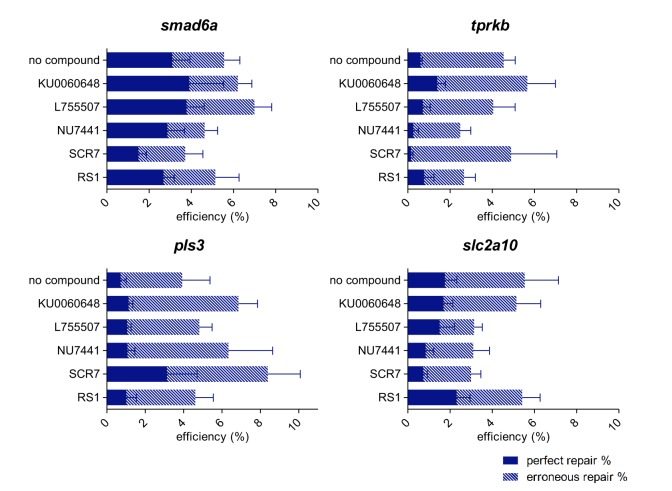
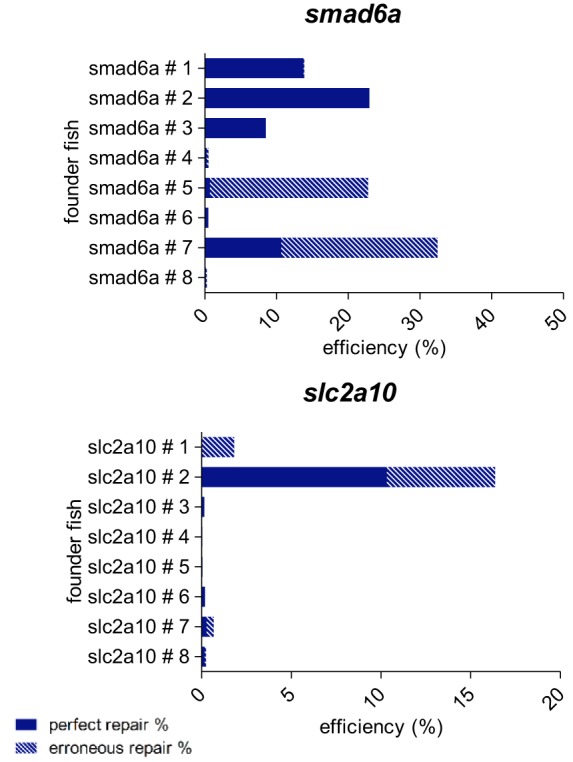
Targeted genome editing by CRISPR/Cas9 is extremely well fitted to generate gene disruptions, although precise sequence replacement by CRISPR/Cas9-mediated homology-directed repair (HDR) suffers from low efficiency, impeding its use for high-throughput knock-in disease modeling. In this study, we used next-generation sequencing (NGS) analysis to determine the efficiency and reliability of CRISPR/Cas9-mediated HDR using several types of single-stranded oligodeoxynucleotide (ssODN) repair templates for the introduction of disease-relevant point mutations in the zebrafish genome. Our results suggest that HDR rates are strongly determined by repair-template composition, with the most influential factor being homology-arm length. However, we found that repair using ssODNs does not only lead to precise sequence replacement but also induces integration of repair-template fragments at the Cas9 cut site. We observed that error-free repair occurs at a relatively constant rate of 1-4% when using different repair templates, which was sufficient for transmission of point mutations to the F1 generation. On the other hand, erroneous repair mainly accounts for the variability in repair rate between the different repair templates. To further improve error-free HDR rates, elucidating the mechanism behind this erroneous repair is essential. We show that the error-prone nature of ssODN-mediated repair, believed to act via synthesis-dependent strand annealing (SDSA), is most likely due to DNA synthesis errors. In conclusion, caution is warranted when using ssODNs for the generation of knock-in models or for therapeutic applications. We recommend the application of in-depth NGS analysis to examine both the efficiency and error-free nature of HDR events.This article has an associated First Person interview with the first author of the paper.
Keywords: CRISPR/Cas9; HDR; Homology-directed repair; Next-generation sequencing; Zebrafish.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
