

Selfish mutations dysregulating RAS-MAPK signaling are pervasive in aged human testes
- PMID: 30355600
- PMCID: PMC6280762
- DOI: 10.1101/gr.239186.118
Selfish mutations dysregulating RAS-MAPK signaling are pervasive in aged human testes
Abstract
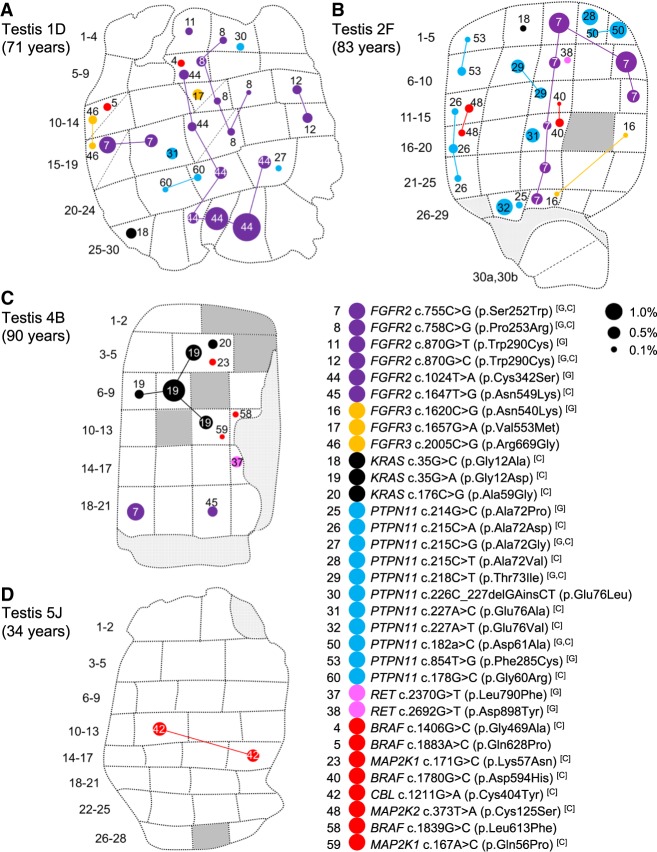
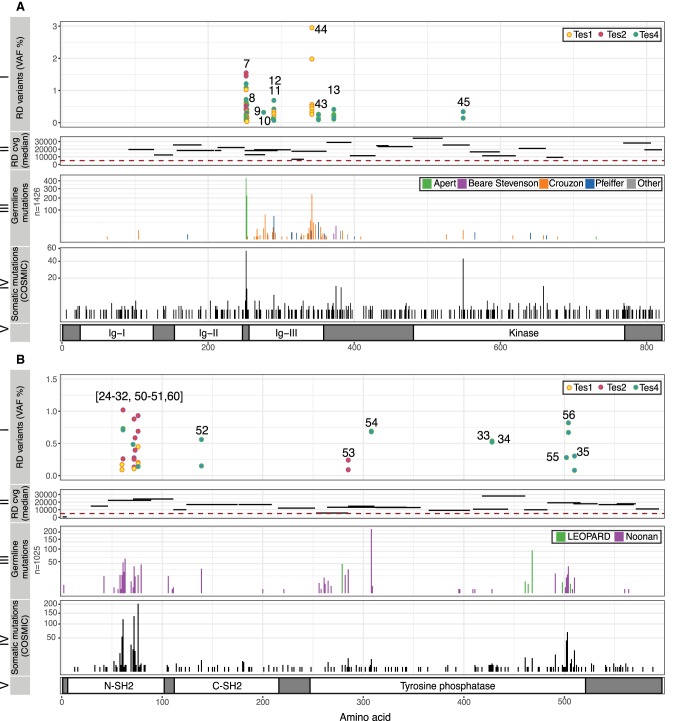
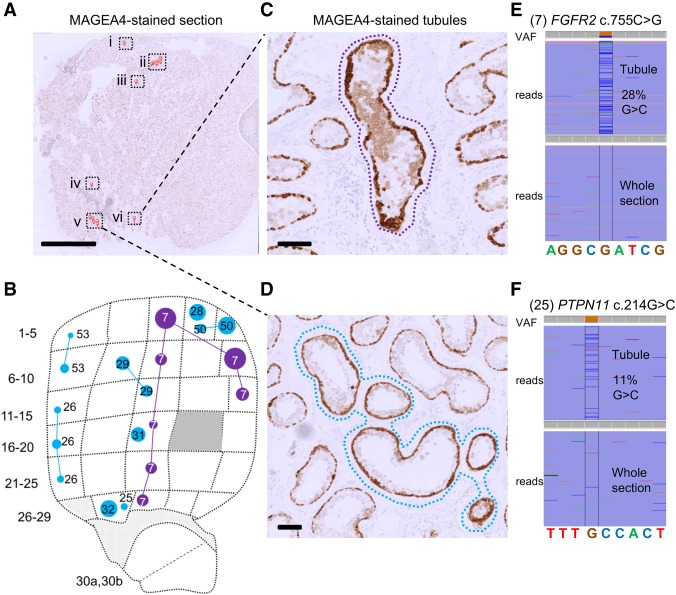
Mosaic mutations present in the germline have important implications for reproductive risk and disease transmission. We previously demonstrated a phenomenon occurring in the male germline, whereby specific mutations arising spontaneously in stem cells (spermatogonia) lead to clonal expansion, resulting in elevated mutation levels in sperm over time. This process, termed "selfish spermatogonial selection," explains the high spontaneous birth prevalence and strong paternal age-effect of disorders such as achondroplasia and Apert, Noonan and Costello syndromes, with direct experimental evidence currently available for specific positions of six genes (FGFR2, FGFR3, RET, PTPN11, HRAS, and KRAS). We present a discovery screen to identify novel mutations and genes showing evidence of positive selection in the male germline, by performing massively parallel simplex PCR using RainDance technology to interrogate mutational hotspots in 67 genes (51.5 kb in total) in 276 biopsies of testes from five men (median age, 83 yr). Following ultradeep sequencing (about 16,000×), development of a low-frequency variant prioritization strategy, and targeted validation, we identified 61 distinct variants present at frequencies as low as 0.06%, including 54 variants not previously directly associated with selfish selection. The majority (80%) of variants identified have previously been implicated in developmental disorders and/or oncogenesis and include mutations in six newly associated genes (BRAF, CBL, MAP2K1, MAP2K2, RAF1, and SOS1), all of which encode components of the RAS-MAPK pathway and activate signaling. Our findings extend the link between mutations dysregulating the RAS-MAPK pathway and selfish selection, and show that the aging male germline is a repository for such deleterious mutations.
© 2018 Maher et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Acuna-Hidalgo R, Sengul H, Steehouwer M, van de Vorst M, Vermeulen SH, Kiemeney L, Veltman JA, Gilissen C, Hoischen A. 2017. Ultra-sensitive sequencing identifies high prevalence of clonal hematopoiesis-associated mutations throughout adult life. Am J Hum Genet 101: 50–64. 10.1016/j.ajhg.2017.05.013 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 091182 /WT_/Wellcome Trust/United Kingdom
- RG/13/13/30194/BHF_/British Heart Foundation/United Kingdom
- 105361 /WT_/Wellcome Trust/United Kingdom
- 102731/WT_/Wellcome Trust/United Kingdom
- MR/L003120/1/MRC_/Medical Research Council/United Kingdom
- 091182/WT_/Wellcome Trust/United Kingdom
- 102731/Z/13/Z/WT_/Wellcome Trust/United Kingdom
- 090532/WT_/Wellcome Trust/United Kingdom
- MC_UU_ 12025/MRC_/Medical Research Council/United Kingdom
- G0902418 /MRC_/Medical Research Council/United Kingdom
- CRUK_/Cancer Research UK/United Kingdom
- 99148/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous