

Eurasian Avian-Like Swine Influenza A Viruses Escape Human MxA Restriction through Distinct Mutations in Their Nucleoprotein
- PMID: 30355693
- PMCID: PMC6321936
- DOI: 10.1128/JVI.00997-18
Eurasian Avian-Like Swine Influenza A Viruses Escape Human MxA Restriction through Distinct Mutations in Their Nucleoprotein
Abstract
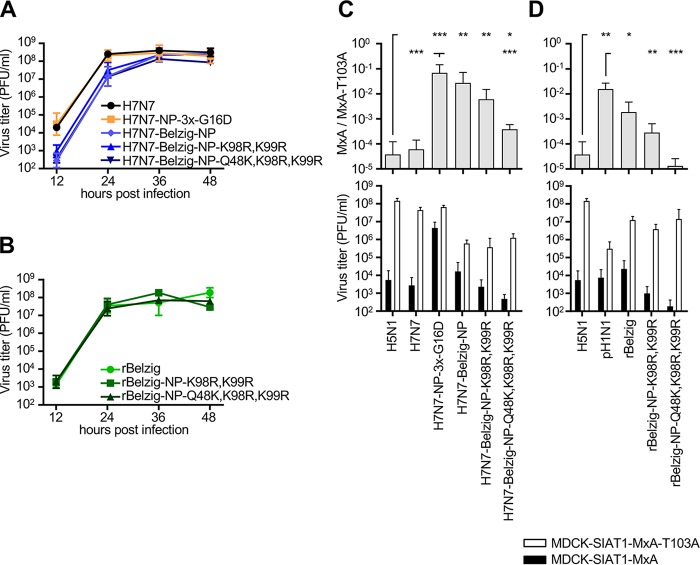
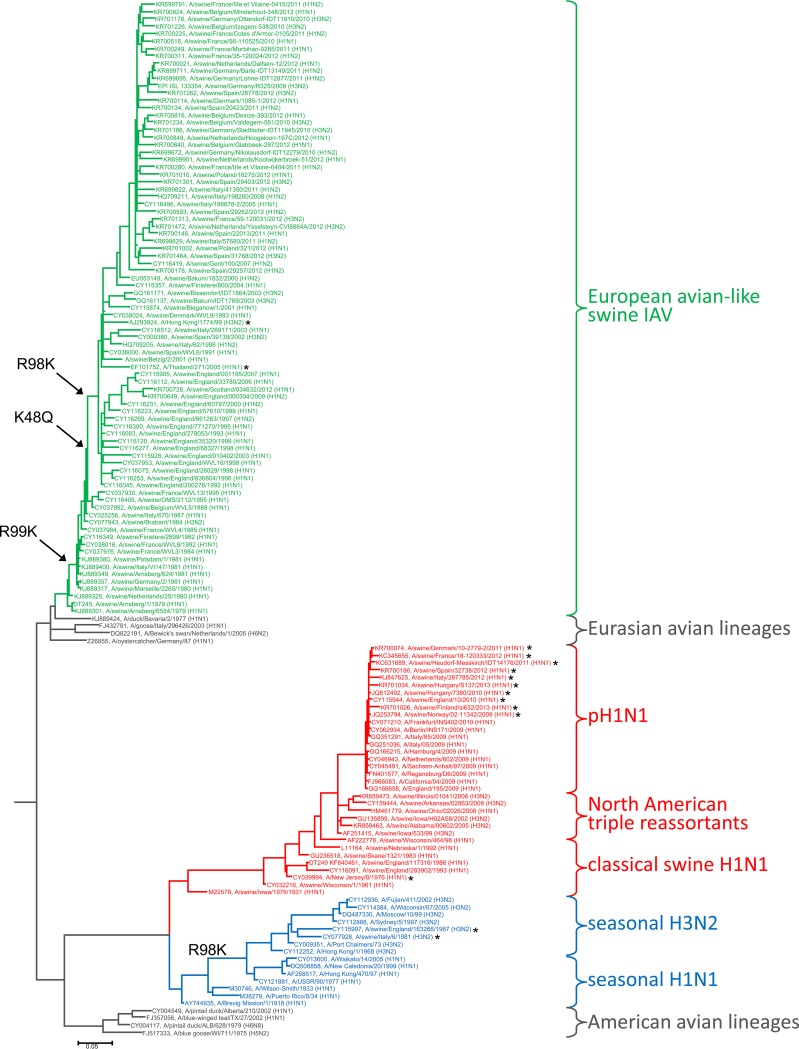
To cross the human species barrier, influenza A viruses (IAV) of avian origin have to overcome the interferon-induced host restriction factor MxA by acquiring distinct mutations in their nucleoprotein (NP). We recently demonstrated that North American classical swine IAV are able to partially escape MxA restriction. Here we investigated whether the Eurasian avian-like swine IAV lineage currently circulating in European swine would likewise evade restriction by human MxA. We found that the NP of the influenza virus isolate A/Swine/Belzig/2/2001 (Belzig-NP) exhibits increased MxA escape, similar in extent to that with human IAV NPs. Mutational analysis revealed that the MxA escape mutations in Belzig-NP differ from the known MxA resistance cluster of the North American classical swine lineage and human-derived IAV NPs. A mouse-adapted avian IAV of the H7N7 subtype encoding Belzig-NP showed significantly greater viral growth in both MxA-expressing cells and MxA-transgenic mice than control viruses lacking the MxA escape mutations. Similarly, the growth of the recombinant Belzig virus was only marginally affected in MxA-expressing cells and MxA-transgenic mice, in contrast to that of Belzig mutant viruses lacking MxA escape mutations in the NP. Phylogenetic analysis of the Eurasian avian-like swine IAV revealed that the NP amino acids required for MxA escape were acquired successively and were maintained after their introduction. Our results suggest that the circulation of IAV in the swine population can result in the selection of NP variants with a high degree of MxA resistance, thereby increasing the zoonotic potential of these viruses. IMPORTANCE The human MxA protein efficiently blocks the replication of IAV from nonhuman species. In rare cases, however, these IAV overcome the species barrier and become pandemic. All known pandemic viruses have acquired and maintained MxA escape mutations in the viral NP and thus are not efficiently controlled by MxA. Intriguingly, partial MxA resistance can also be acquired in other hosts that express antivirally active Mx proteins, such as swine. To perform a risk assessment of IAV circulating in the European swine population, we analyzed the degree of MxA resistance of Eurasian avian-like swine IAV. Our data demonstrate that these viruses carry formerly undescribed Mx resistance mutations in the NP that mediate efficient escape from human MxA. We conclude that Eurasian avian-like swine IAV possess substantial zoonotic potential.
Copyright © 2018 American Society for Microbiology.
Figures
References
-
- Vincent A, Awada L, Brown I, Chen H, Claes F, Dauphin G, Donis R, Culhane M, Hamilton K, Lewis N, Mumford E, Nguyen T, Parchariyanon S, Pasick J, Pavade G, Pereda A, Peiris M, Saito T, Swenson S, Van Reeth K, Webby R, Wong F, Ciacci-Zanella J. 2014. Review of influenza A virus in swine worldwide: a call for increased surveillance and research. Zoonoses Public Health 61:4–17. doi: 10.1111/zph.12049. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
