

Size matters: height, cell number and a person's risk of cancer
- PMID: 30355711
- PMCID: PMC6234890
- DOI: 10.1098/rspb.2018.1743
Size matters: height, cell number and a person's risk of cancer
Abstract
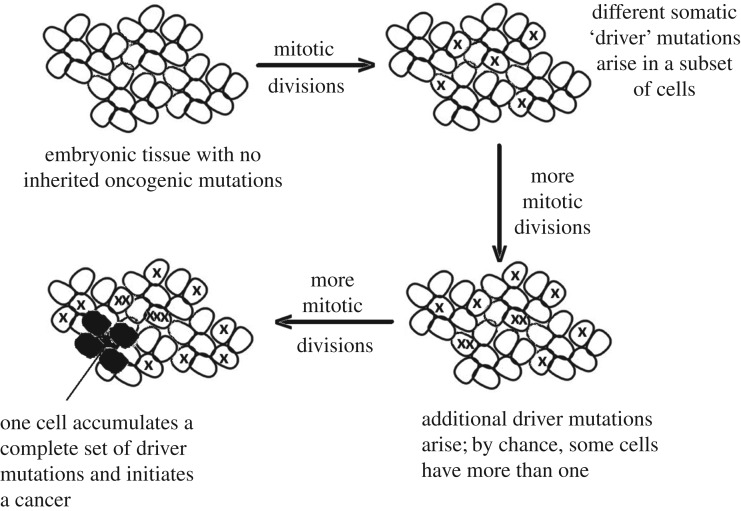
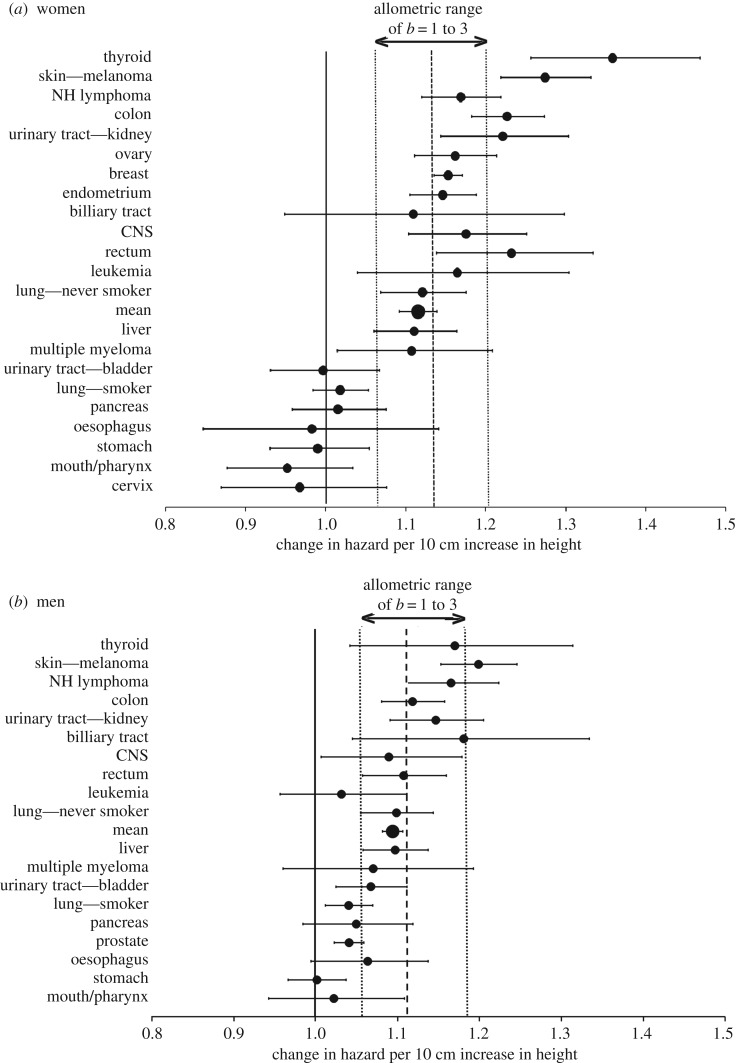
The multistage model of carcinogenesis predicts cancer risk will increase with tissue size, since more cells provide more targets for oncogenic somatic mutation. However, this increase is not seen among mammal species of different sizes (Peto's paradox), a paradox argued to be due to larger species evolving added cancer suppression. If this explanation is correct, the cell number effect is still expected within species. Consistent with this, the hazard ratio for overall cancer risk per 10 cm increase in human height (HR10) is about 1.1, indicating a 10% increase in cancer risk per 10 cm; however, an alternative explanation invokes an indirect effect of height, with factors that increase cancer risk independently increasing adult height. The data from four large-scale surveillance projects on 23 cancer categories were tested against quantitative predictions of the cell-number hypothesis, predictions that were accurately supported. For overall cancer risk the HR10 predicted versus observed was 1.13 versus 1.12 for women and 1.11 versus 1.09 for men, suggesting that cell number variation provides a null hypothesis for assessing height effects. Melanoma showed an unexpectedly strong relationship to height, indicating an additional effect, perhaps due to an increasing cell division rate mediated through increasing IGF-I with height. Similarly, only about one-third of the higher incidence of non-reproductive cancers in men versus women can be explained by cell number. The cancer risks of obesity are not correlated with effects of height, consistent with different primary causation. The direct effect of height on cancer risk suggests caution in identifying height-related SNPs as cancer causing.
Keywords: GWAS and cancer; Peto's paradox; cancer and BMI; cancer and height; cell number; multistage carcinogenesis.
© 2018 The Author(s).
Conflict of interest statement
I have no competing interests.
Figures
References
-
- Peto R. 1977. Epidemiology, multistage models, and short-term mutagenicity tests. In The origins of human cancer, vol. 4 (eds Hiatt HH, Watson JD, Winsten JA), pp. 1403–1428. New York, NY: Cold Spring Harbor Laboratory Press.
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources