

Coaching from the sidelines: the nuclear periphery in genome regulation
- PMID: 30356165
- PMCID: PMC6355253
- DOI: 10.1038/s41576-018-0063-5
Coaching from the sidelines: the nuclear periphery in genome regulation
Abstract
The genome is packaged and organized nonrandomly within the 3D space of the nucleus to promote efficient gene expression and to faithfully maintain silencing of heterochromatin. The genome is enclosed within the nucleus by the nuclear envelope membrane, which contains a set of proteins that actively participate in chromatin organization and gene regulation. Technological advances are providing views of genome organization at unprecedented resolution and are beginning to reveal the ways that cells co-opt the structures of the nuclear periphery for nuclear organization and gene regulation. These genome regulatory roles of proteins of the nuclear periphery have important influences on development, disease and ageing.
Figures


References
-
- Cremer T & Cremer C Rise, fall and resurrection of chromosome territories: a historical perspective. Part I. The rise of chromosome territories. Eur. J. Histochem 50, 161–176 (2006). - PubMed
-
- Sati S & Cavalli G Chromosome conformation capture technologies and their impact in understanding genome function. Chromosoma 126, 33–44 (2017). - PubMed
-
- Lieberman-Aiden E et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 326, 289–293 (2009). - PMC - PubMed
-
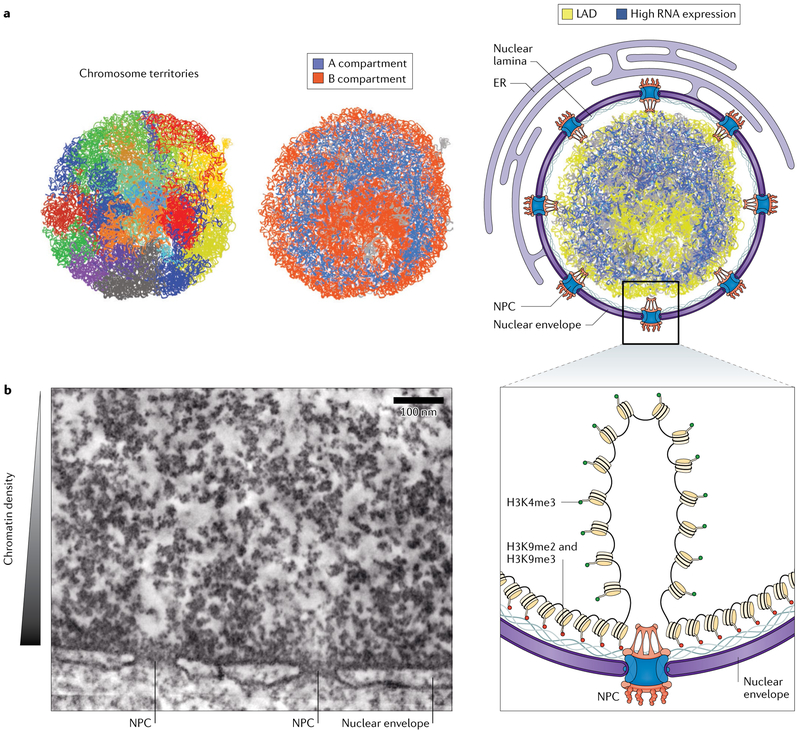
This article provides the first demonstration of the HiC method for probing genome conformation indicating global properties of chromatin folding — that chromosomes occupy distinct territories and that chromatin separates into megabase-scale A and B compartments on the basis of chromatin activity and gene density.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
