

Nesting Environment Provides Sex-Specific Neuroprotection in a Rat Model of Neonatal Hypoxic-Ischemic Injury
- PMID: 30356904
- PMCID: PMC6190890
- DOI: 10.3389/fnbeh.2018.00221
Nesting Environment Provides Sex-Specific Neuroprotection in a Rat Model of Neonatal Hypoxic-Ischemic Injury
Abstract
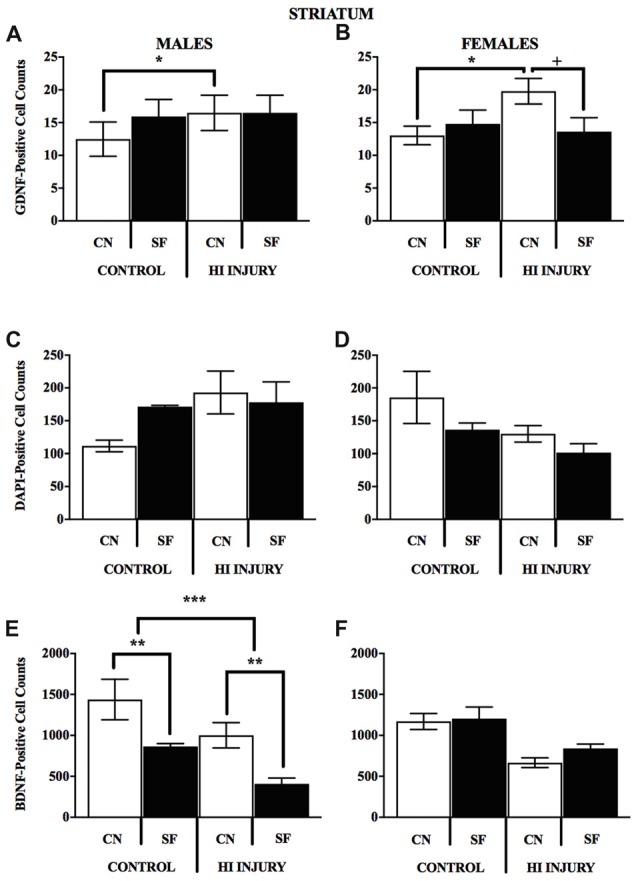
Hypoxic-ischemic (HI) encephalopathy is a devastating injury that occurs when the fetal brain is deprived of oxygen and blood to a degree that may lead to neurological damage, seizing and cerebral palsy. In rodents, early environmental enrichment that promotes maternal care-taking behavior (mCTB) can improve neurobehavioral outcomes and protect against neurological decline. We hypothesized that an enhanced nesting environment would improve mCTB as measured by pup weight gain, and support greater HI recovery in developing rats. Pregnant dams (E15-16) were introduced to either control Standard Facility (SF) housing or closed nestbox (CN) conditions and maintained in larger cages through pup weaning. On postnatal day (PND) 7, male and female Long-Evans rat pups (N = 73) were randomly sorted into one of two surgical conditions: control and HI. HI pups received isoflurane anesthesia and right carotid artery ligation, a 2-h rest followed by 90 min exposure to a moist hypoxic (92% N, 8% O2) chamber. Pups (PND 8) were weighed daily, and tested on the Morris Water Maze (MWM) task (PND 35-50). Results demonstrate significant differences afforded to male and female pups based on weight measure, where CN-rearing modifies pre-weaning adolescent weights in females and increases post-weaning weights in males and females by an average of 10 g. Following successful MWM training and acquisition (PND 35-37), both male and female CN-raised animals demonstrated faster latency to find the hidden platform (HP) during HP trials (PND 38-42) and appeared to freely explore the MWM pool during an additional probe trial (PND 43). Moreover, after sacrifice (PND 60), CN rearing created sex-specific alterations in brain-derived neurotrophic factor (BDNF), glial-derived neurotrophic factor (GDNF) immunopositive cell staining of the dorsomedial striatum and CA1 of the hippocampus. CN-rearing afforded HI males higher BDNF levels in the striatum and produced greater GDNF levels in the hippocampus of HI-injured females. These results suggest that early life environmental enrichment positively modifies nesting environment, increases weight gain, as well as spatial learning and memory in a sex-specific directionality. Our findings also implicate correlative changes in corticolimbic neurotrophin protein levels in the CN-reared animals that may contribute to these benefits.
Keywords: Long Evans rats; Morris water maze; Rice-Vanucci P7 HI model; environmental enrichment; hippocampus; neonatal hypoxic ischemic injury; neurogenesis.
Figures









Similar articles
-
Severe Perinatal Hypoxic-Ischemic Brain Injury Induces Long-Term Sensorimotor Deficits, Anxiety-Like Behaviors and Cognitive Impairment in a Sex-, Age- and Task-Selective Manner in C57BL/6 Mice but Can Be Modulated by Neonatal Handling.Front Behav Neurosci. 2019 Feb 13;13:7. doi: 10.3389/fnbeh.2019.00007. eCollection 2019. Front Behav Neurosci. 2019. PMID: 30814939 Free PMC article.
-
Environmental enrichment attenuates the blood brain barrier dysfunction induced by the neonatal hypoxia-ischemia.Int J Dev Neurosci. 2016 Oct;53:35-45. doi: 10.1016/j.ijdevneu.2016.06.006. Epub 2016 Jun 17. Int J Dev Neurosci. 2016. PMID: 27326908
-
NTP technical report on the toxicity studies of Dibutyl Phthalate (CAS No. 84-74-2) Administered in Feed to F344/N Rats and B6C3F1 Mice.Toxic Rep Ser. 1995 Apr;30:1-G5. Toxic Rep Ser. 1995. PMID: 12209194
-
Interaction does Count: A Cross-Fostering Study on Transgenerational Effects of Pre-reproductive Maternal Enrichment.Front Behav Neurosci. 2015 Dec 1;9:320. doi: 10.3389/fnbeh.2015.00320. eCollection 2015. Front Behav Neurosci. 2015. PMID: 26648854 Free PMC article.
-
Potential biomarkers for neuroinflammation and neurodegeneration at short and long term after neonatal hypoxic-ischemic insult in rat.J Neuroinflammation. 2019 Oct 28;16(1):194. doi: 10.1186/s12974-019-1595-0. J Neuroinflammation. 2019. PMID: 31660990 Free PMC article.
Cited by
-
CXCR4 inhibition with AMD3100 attenuates amphetamine induced locomotor activity in adolescent Long Evans male rats.PLoS One. 2021 Mar 1;16(3):e0247707. doi: 10.1371/journal.pone.0247707. eCollection 2021. PLoS One. 2021. PMID: 33647040 Free PMC article.
-
Oligodendrocyte progenitor cells' fate after neonatal asphyxia-Puzzling implications for the development of hypoxic-ischemic encephalopathy.Brain Pathol. 2024 Nov;34(6):e13255. doi: 10.1111/bpa.13255. Epub 2024 Mar 19. Brain Pathol. 2024. PMID: 38504469 Free PMC article.
-
Effective Hand Rearing of Neonatal Mice for Developmental Studies.Bio Protoc. 2023 Jun 5;13(11):e4755. doi: 10.21769/BioProtoc.4755. eCollection 2023 Jun 5. Bio Protoc. 2023. PMID: 37323632 Free PMC article.
-
Environmental enrichment with nylon gnaw sticks introduces variation in Sprague Dawley rat immune and lower gastrointestinal parameters with differences between sexes.Anim Microbiome. 2025 Jan 31;7(1):12. doi: 10.1186/s42523-024-00369-6. Anim Microbiome. 2025. PMID: 39891232 Free PMC article.
-
Severe Perinatal Hypoxic-Ischemic Brain Injury Induces Long-Term Sensorimotor Deficits, Anxiety-Like Behaviors and Cognitive Impairment in a Sex-, Age- and Task-Selective Manner in C57BL/6 Mice but Can Be Modulated by Neonatal Handling.Front Behav Neurosci. 2019 Feb 13;13:7. doi: 10.3389/fnbeh.2019.00007. eCollection 2019. Front Behav Neurosci. 2019. PMID: 30814939 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous