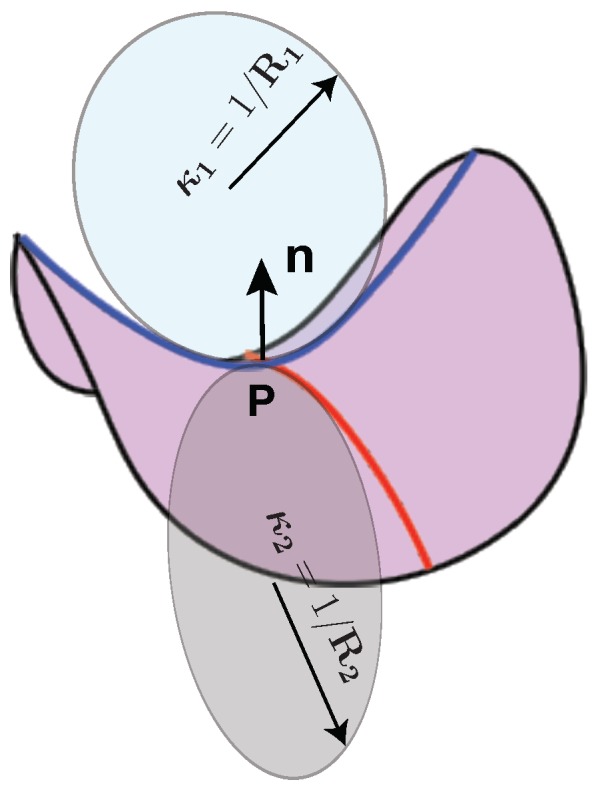
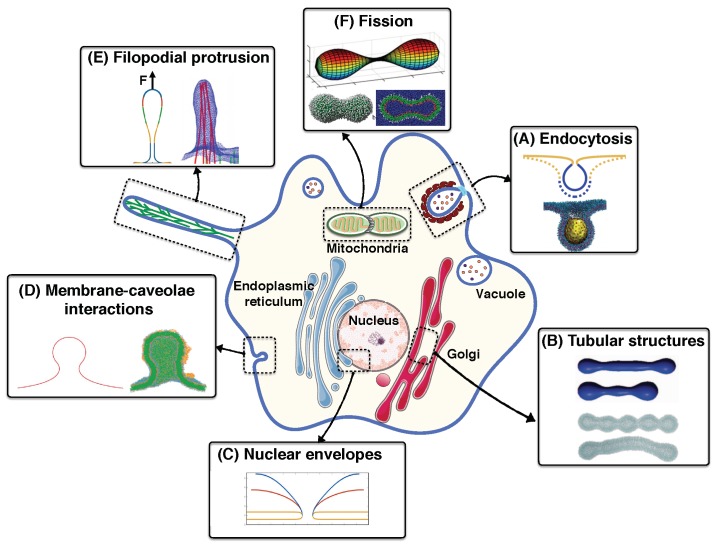
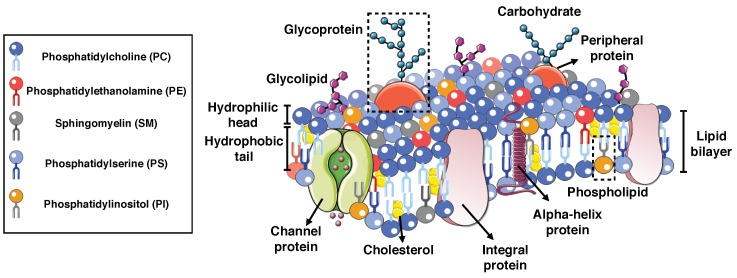
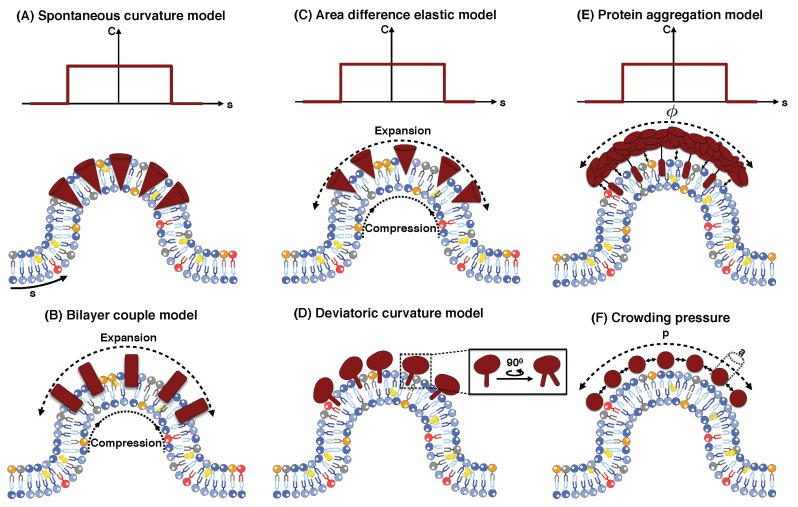
Modeling Membrane Curvature Generation due to Membrane⁻Protein Interactions
- PMID: 30360496
- PMCID: PMC6316661
- DOI: 10.3390/biom8040120
Modeling Membrane Curvature Generation due to Membrane⁻Protein Interactions
Abstract
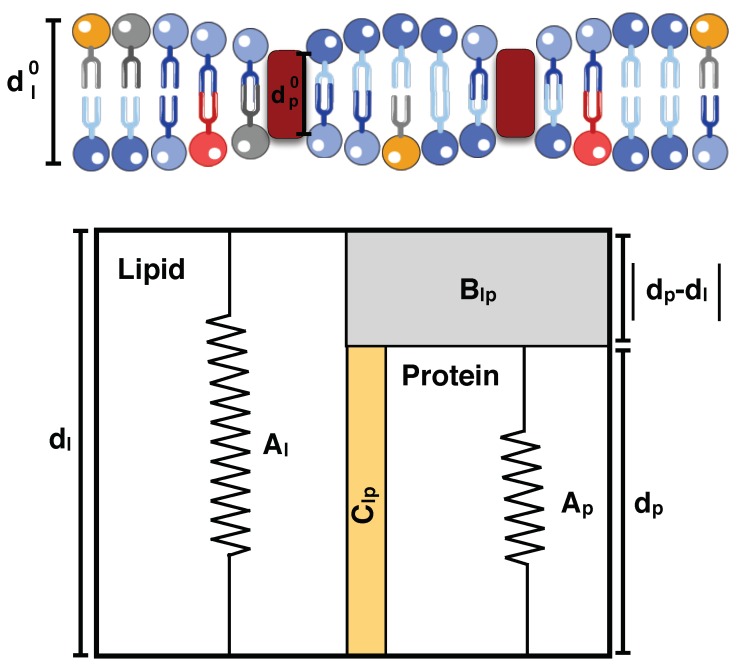
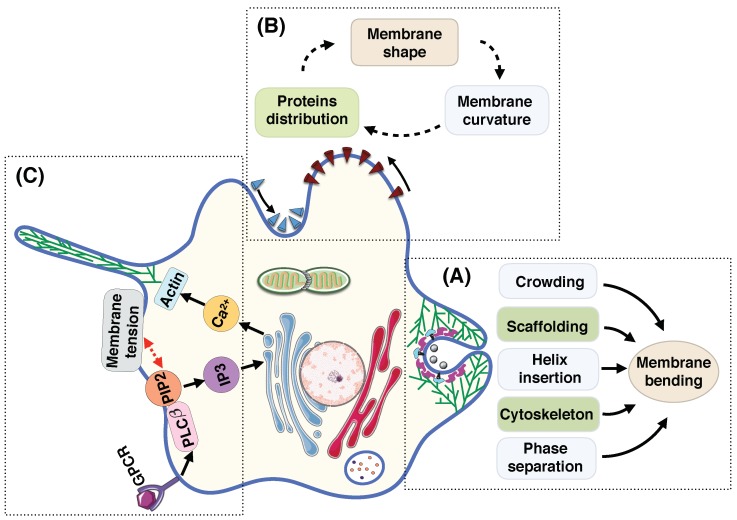
To alter and adjust the shape of the plasma membrane, cells harness various mechanisms of curvature generation. Many of these curvature generation mechanisms rely on the interactions between peripheral membrane proteins, integral membrane proteins, and lipids in the bilayer membrane. Mathematical and computational modeling of membrane curvature generation has provided great insights into the physics underlying these processes. However, one of the challenges in modeling these processes is identifying the suitable constitutive relationships that describe the membrane free energy including protein distribution and curvature generation capability. Here, we review some of the commonly used continuum elastic membrane models that have been developed for this purpose and discuss their applications. Finally, we address some fundamental challenges that future theoretical methods need to overcome to push the boundaries of current model applications.
Keywords: Helfrich energy; area difference elastic model; deviatoric curvature; hydrophobic mismatch; plasma membrane; protein crowding; spontaneous curvature.
Conflict of interest statement
The authors declare no conflict of interest.
Figures