

Pathogen blockade of TAK1 triggers caspase-8-dependent cleavage of gasdermin D and cell death
- PMID: 30361383
- PMCID: PMC6522129
- DOI: 10.1126/science.aau2818
Pathogen blockade of TAK1 triggers caspase-8-dependent cleavage of gasdermin D and cell death
Abstract
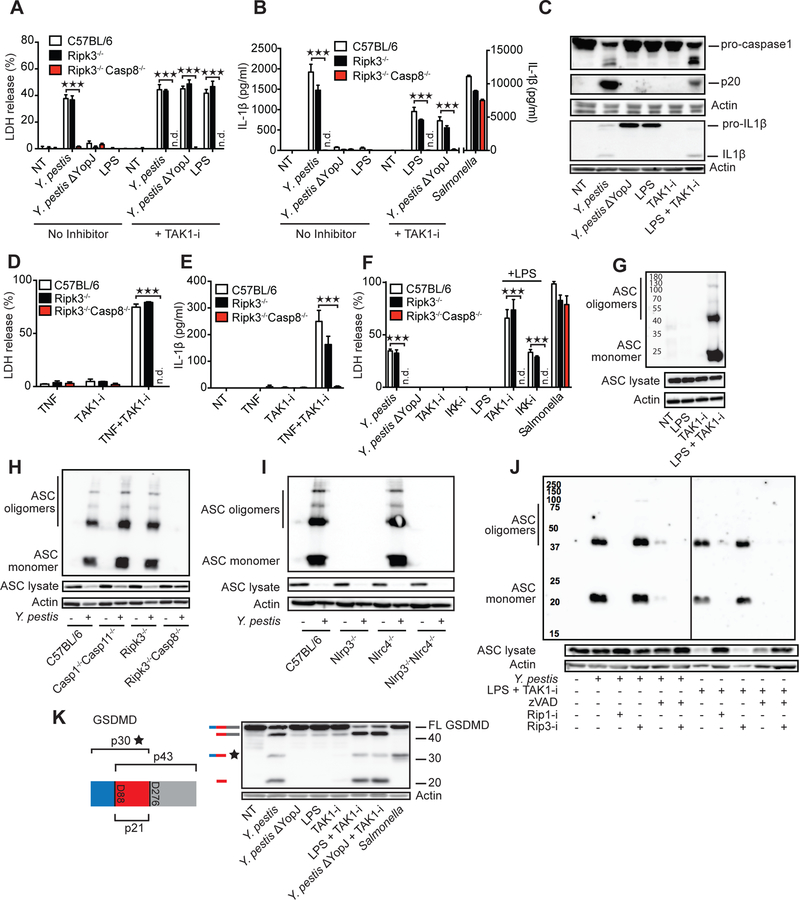
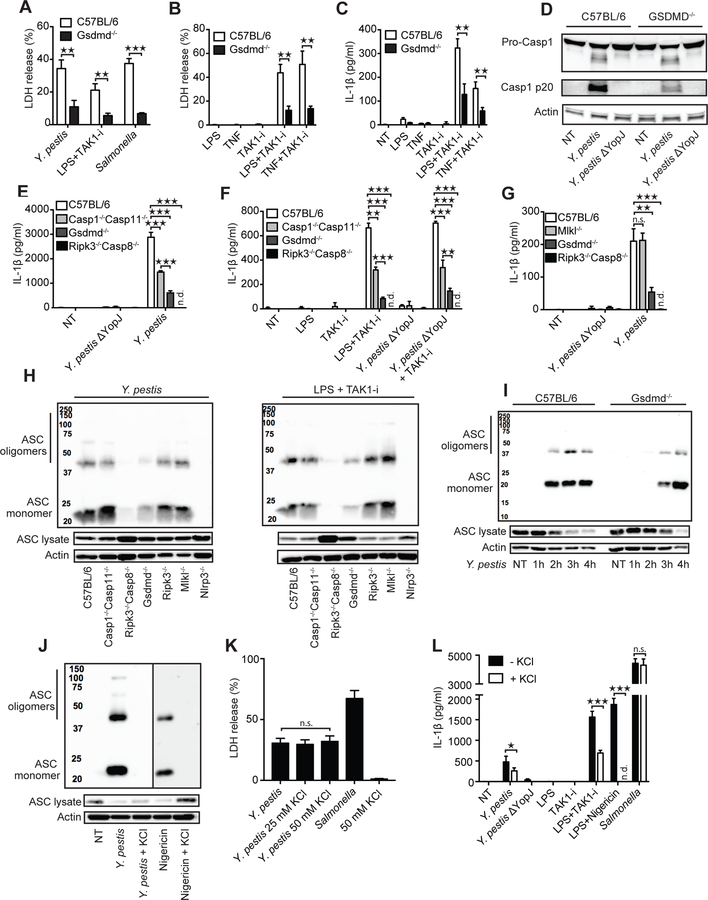
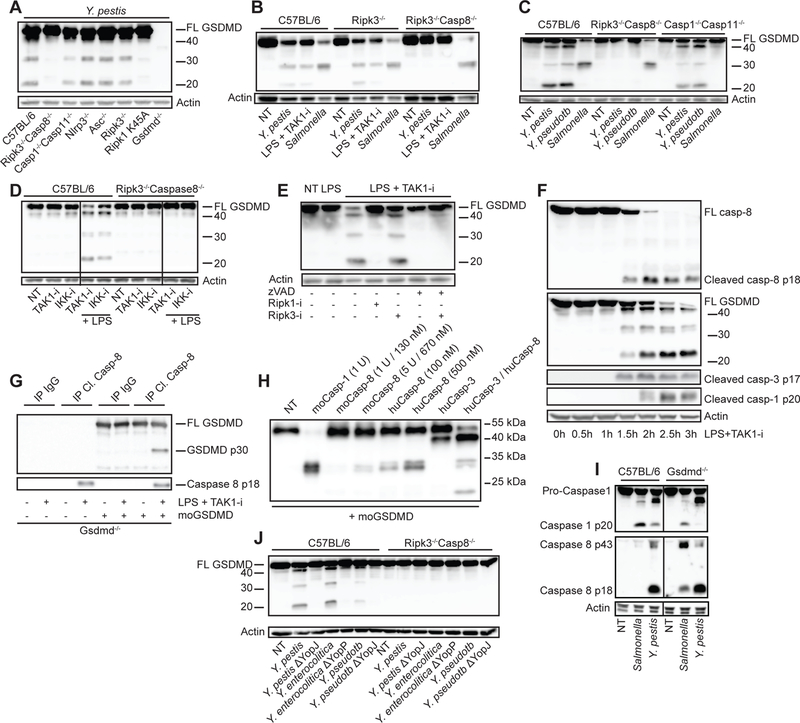
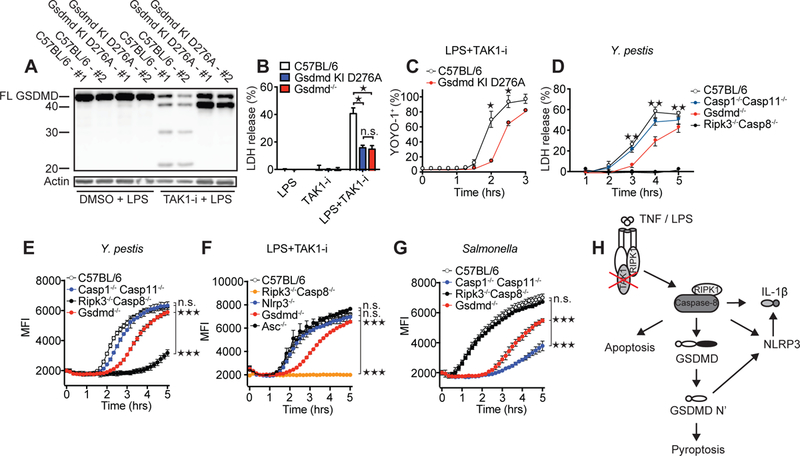
Limited proteolysis of gasdermin D (GSDMD) generates an N-terminal pore-forming fragment that controls pyroptosis in macrophages. GSDMD is processed via inflammasome-activated caspase-1 or -11. It is currently unknown whether macrophage GSDMD can be processed by other mechanisms. Here, we describe an additional pathway controlling GSDMD processing. The inhibition of TAK1 or IκB kinase (IKK) by the Yersinia effector protein YopJ elicits RIPK1- and caspase-8-dependent cleavage of GSDMD, which subsequently results in cell death. GSDMD processing also contributes to the NLRP3 inflammasome-dependent release of interleukin-1β (IL-1β). Thus, caspase-8 acts as a regulator of GSDMD-driven cell death. Furthermore, this study establishes the importance of TAK1 and IKK activity in the control of GSDMD cleavage and cytotoxicity.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI075318/AI/NIAID NIH HHS/United States
- R21 AI128547/AI/NIAID NIH HHS/United States
- R25 HL092610/HL/NHLBI NIH HHS/United States
- R56 AI075118/AI/NIAID NIH HHS/United States
- T32 AI007538/AI/NIAID NIH HHS/United States
- T32 AI095213/AI/NIAID NIH HHS/United States
- DP1 HD087988/HD/NICHD NIH HHS/United States
- R01 AI075118/AI/NIAID NIH HHS/United States
- R37 AI067497/AI/NIAID NIH HHS/United States
- R56 AI067497/AI/NIAID NIH HHS/United States
- R01 AI124491/AI/NIAID NIH HHS/United States
- F30 AI129237/AI/NIAID NIH HHS/United States
- R01 AI139914/AI/NIAID NIH HHS/United States
- R01 AI067497/AI/NIAID NIH HHS/United States
- R01 AI083713/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
