

Adhesion to nanofibers drives cell membrane remodeling through one-dimensional wetting
- PMID: 30361638
- PMCID: PMC6202395
- DOI: 10.1038/s41467-018-06948-x
Adhesion to nanofibers drives cell membrane remodeling through one-dimensional wetting
Abstract
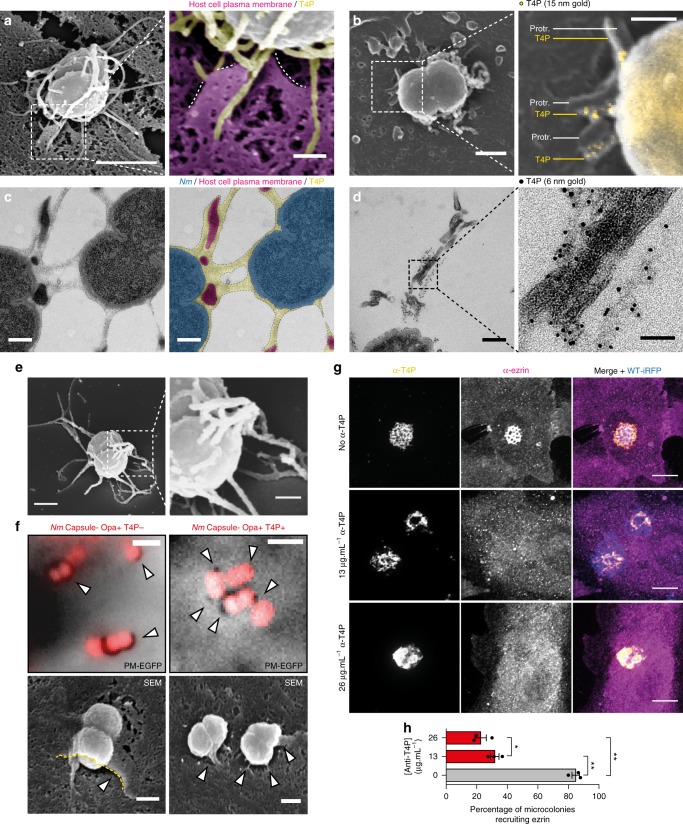
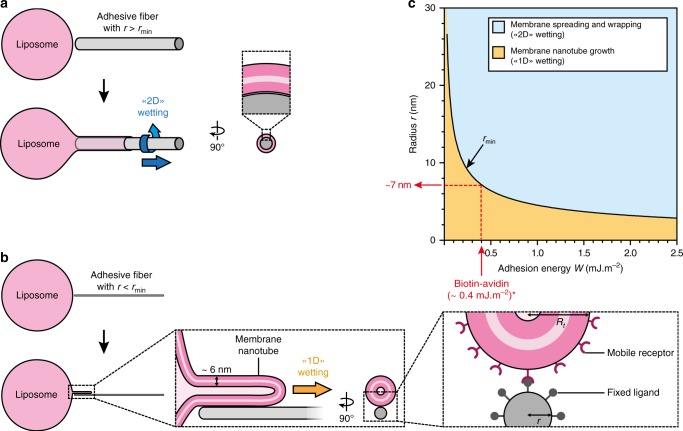
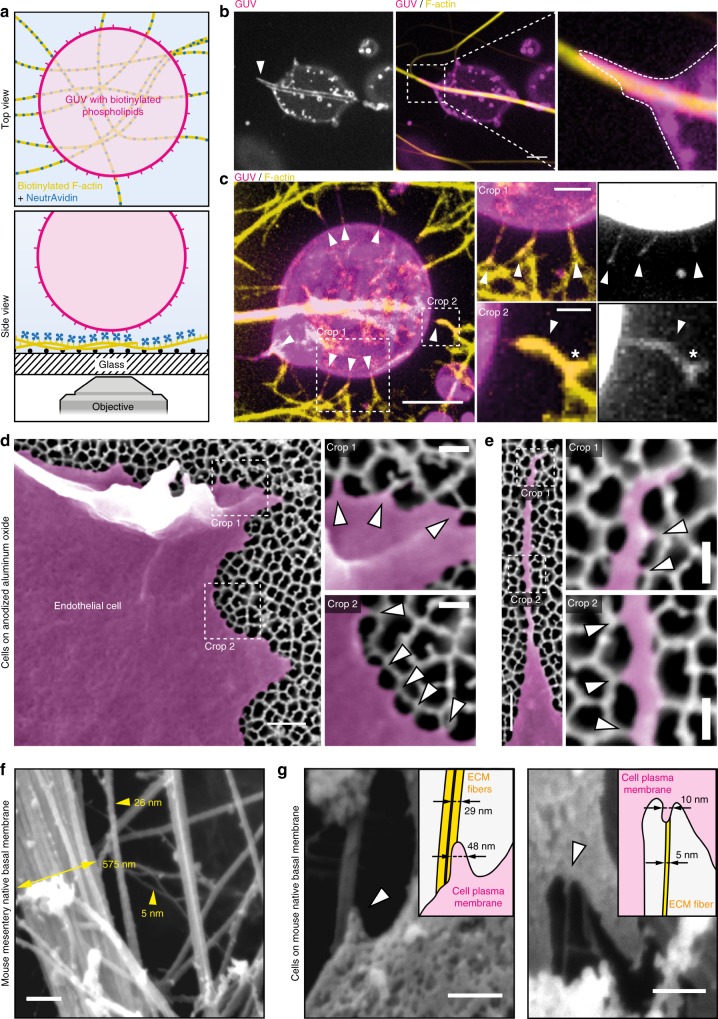
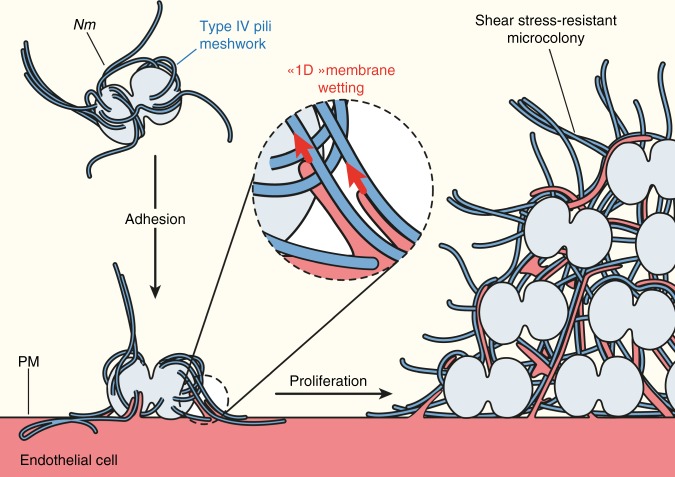
The shape of cellular membranes is highly regulated by a set of conserved mechanisms that can be manipulated by bacterial pathogens to infect cells. Remodeling of the plasma membrane of endothelial cells by the bacterium Neisseria meningitidis is thought to be essential during the blood phase of meningococcal infection, but the underlying mechanisms are unclear. Here we show that plasma membrane remodeling occurs independently of F-actin, along meningococcal type IV pili fibers, by a physical mechanism that we term 'one-dimensional' membrane wetting. We provide a theoretical model that describes the physical basis of one-dimensional wetting and show that this mechanism occurs in model membranes interacting with nanofibers, and in human cells interacting with extracellular matrix meshworks. We propose one-dimensional wetting as a new general principle driving the interaction of cells with their environment at the nanoscale that is diverted by meningococci during infection.
Conflict of interest statement
The authors declare no competing interests.
Figures

Comment in
-
Nanofibres induce remodelling of cell membranes.Nature. 2018 Nov;563(7732):481-482. doi: 10.1038/d41586-018-07261-9. Nature. 2018. PMID: 30459369 No abstract available.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
