

Characterization and regulation of wild-type and mutant TASK-1 two pore domain potassium channels indicated in pulmonary arterial hypertension
- PMID: 30365877
- PMCID: PMC6376074
- DOI: 10.1113/JP277275
Characterization and regulation of wild-type and mutant TASK-1 two pore domain potassium channels indicated in pulmonary arterial hypertension
Abstract
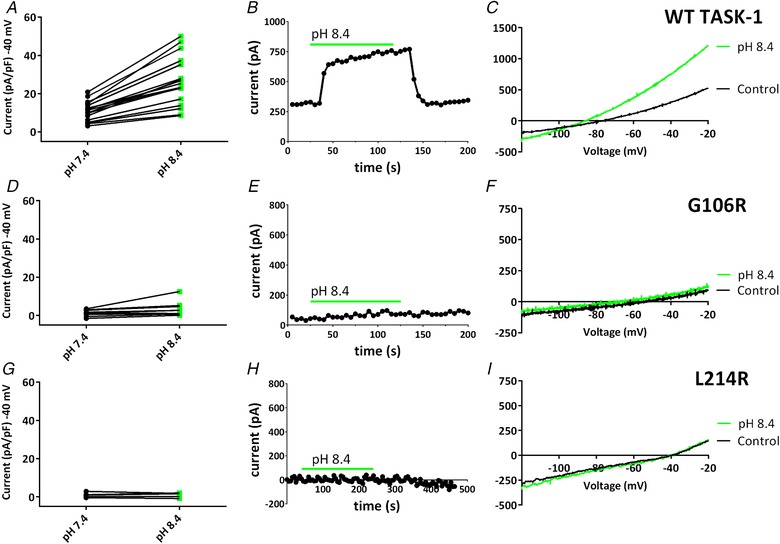
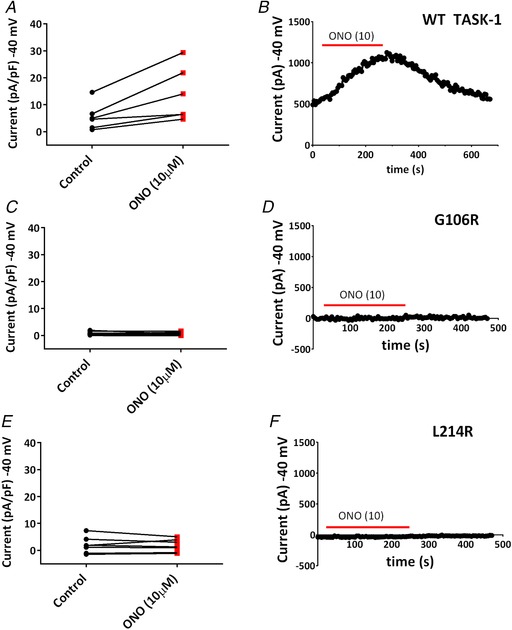
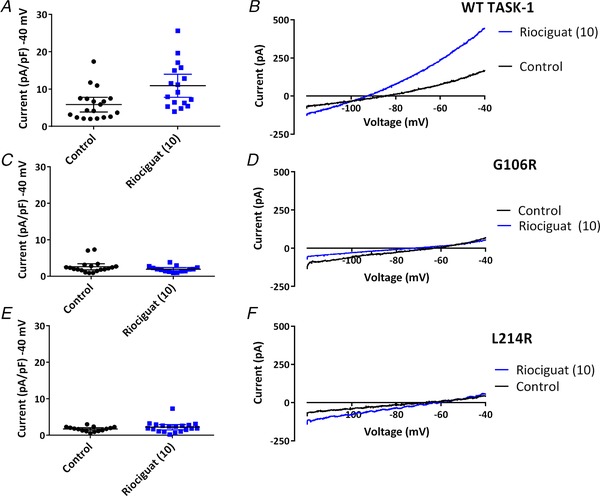
Key points: The TASK-1 channel gene (KCNK3) has been identified as a possible disease-causing gene in heritable pulmonary arterial hypertension (PAH). In the present study, we show that novel mutated TASK-1 channels, seen in PAH patients, have a substantially reduced current compared to wild-type TASK-1 channels. These mutated TASK-1 channels are located at the plasma membrane to the same degree as wild-type TASK-1 channels. ONO-RS-082 and alkaline pH 8.4 both activate TASK-1 channels but do not recover current through mutant TASK-1 channels. We show that the guanylate cyclase activator, riociguat, a novel treatment for PAH, enhances current through TASK-1 channels but does not recover current through mutant TASK-1 channels.
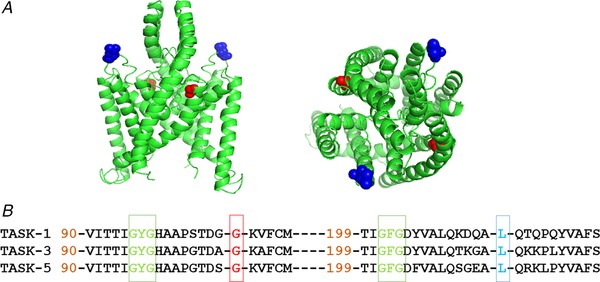
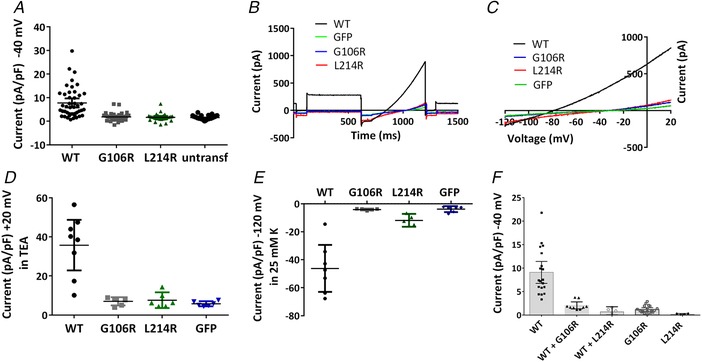
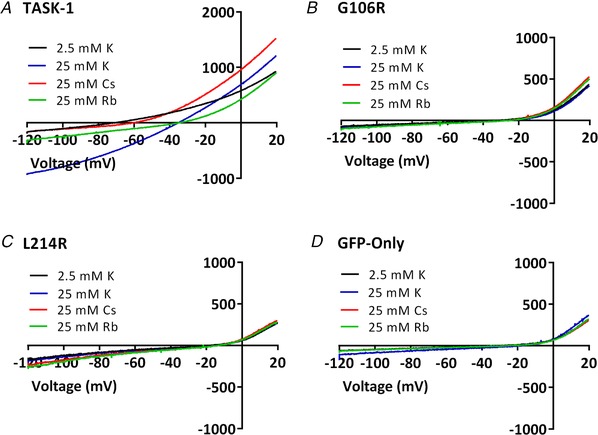
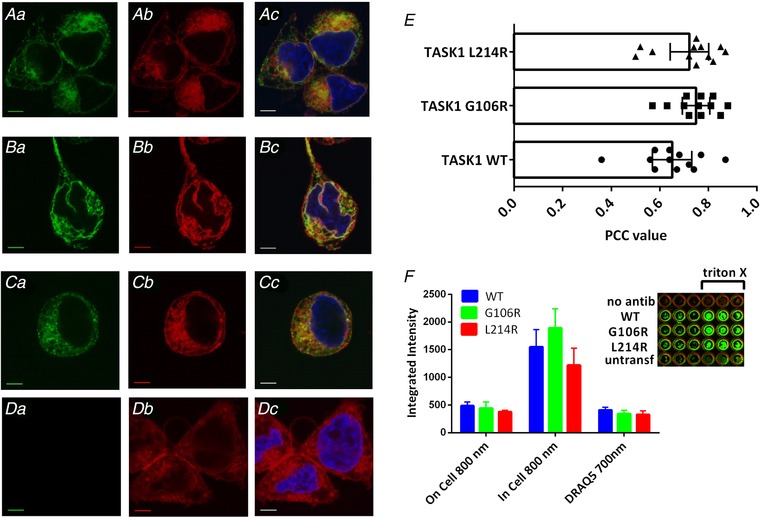
Abstract: Pulmonary arterial hypertension (PAH) affects ∼15-50 people per million. KCNK3, the gene that encodes the two pore domain potassium channel TASK-1 (K2P3.1), has been identified as a possible disease-causing gene in heritable PAH. Recently, two new mutations have been identified in KCNK3 in PAH patients: G106R and L214R. The present study aimed to characterize the functional properties and regulation of wild-type (WT) and mutated TASK-1 channels and determine how these might contribute to PAH and its treatment. Currents through WT and mutated human TASK-1 channels transiently expressed in tsA201 cells were measured using whole-cell patch clamp electrophysiology. Localization of fluorescence-tagged channels was visualized using confocal microscopy and quantified with in-cell and on-cell westerns. G106R or L214R mutated channels were located at the plasma membrane to the same degree as WT channels; however, their current was markedly reduced compared to WT TASK-1 channels. Functional current through these mutated channels could not be restored using activators of WT TASK-1 channels (pH 8.4, ONO-RS-082). The guanylate cyclase activator, riociguat, enhanced current through WT TASK-1 channels; however, similar to the other activators investigated, riociguat did not have any effect on current through mutated TASK-1 channels. Thus, novel mutations in TASK-1 seen in PAH substantially alter the functional properties of these channels. Current through these channels could not be restored by activators of TASK-1 channels. Riociguat enhancement of current through TASK-1 channels could contribute to its therapeutic benefit in the treatment of PAH.
Keywords: KCNK3 (TASK-1) potassium channel; Pulmonary arterial hypertension; riociguat.
© 2018 The Authors. The Journal of Physiology published by John Wiley & Sons Ltd on behalf of The Physiological Society.
Figures
References
-
- Antigny F, Hautefort A, Meloche J, Belacel‐Ouari M, Manoury B, Rucker‐Martin C, Péchoux C, Potus F, Nadeau V, Tremblay E, Ruffenach G, Bourgeois A, Dorfmüller P, Breuils‐Bonnet S, Fadel E, Ranchoux B, Jourdon P, Girerd B, Montani D, Provencher S, Bonnet S, Simonneau G, Humbert M & Perros F (2016). Potassium channel subfamily K member 3 (KCNK3) contributes to the development of pulmonary arterial hypertension. Circulation 133, 1371–1385. - PubMed
-
- Boucherat O, Chabot S, Antigny F, Perros F, Provencher S & Bonnet S (2015). Potassium channels in pulmonary arterial hypertension. Eur Respir J 46, 1167–1177. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical