

LMI1 homeodomain protein regulates organ proportions by spatial modulation of endoreduplication
- PMID: 30366902
- PMCID: PMC6217736
- DOI: 10.1101/gad.318212.118
LMI1 homeodomain protein regulates organ proportions by spatial modulation of endoreduplication
Erratum in
-
Corrigendum: LMI1 homeodomain protein regulates organ proportions by spatial modulation of endoreduplication.Genes Dev. 2019 Mar 1;33(5-6):377. doi: 10.1101/gad.323733.118. Genes Dev. 2019. PMID: 30824533 Free PMC article. No abstract available.
Abstract
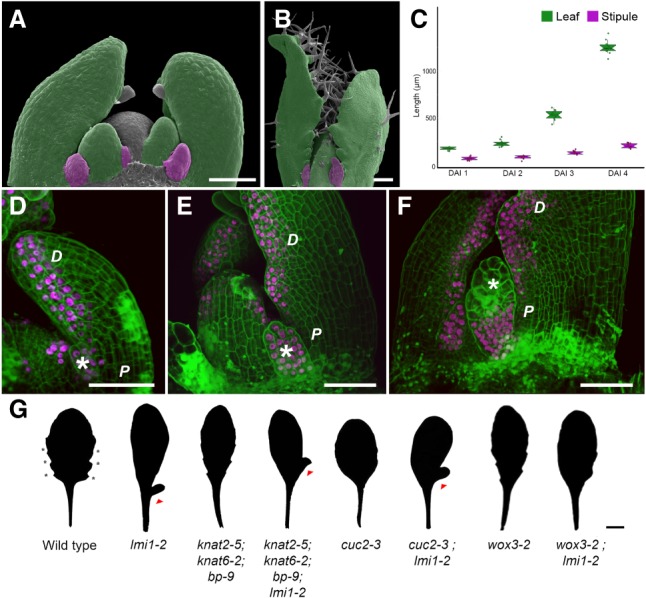
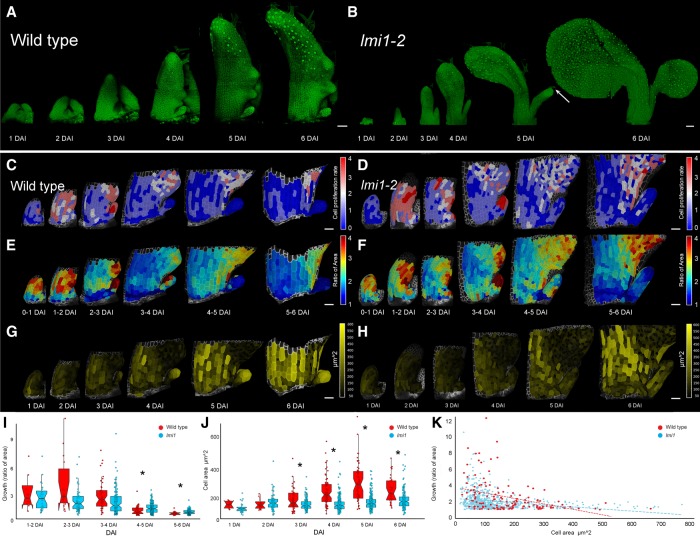
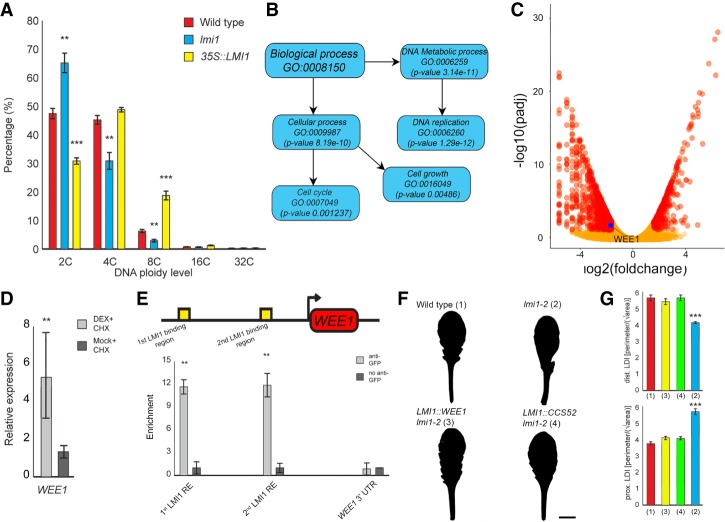
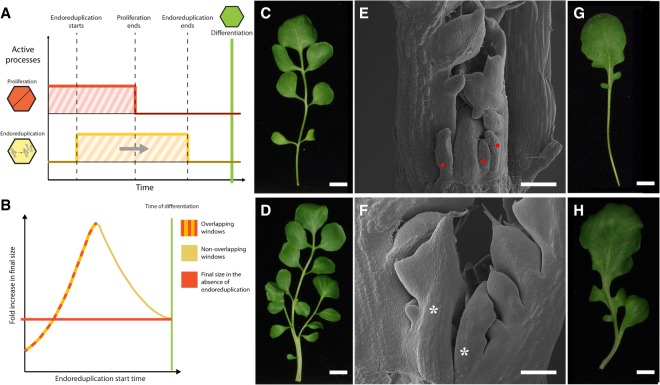
How the interplay between cell- and tissue-level processes produces correctly proportioned organs is a key problem in biology. In plants, the relative size of leaves compared with their lateral appendages, called stipules, varies tremendously throughout development and evolution, yet relevant mechanisms remain unknown. Here we use genetics, live imaging, and modeling to show that in Arabidopsis leaves, the LATE MERISTEM IDENTITY1 (LMI1) homeodomain protein regulates stipule proportions via an endoreduplication-dependent trade-off that limits tissue size despite increasing cell growth. LM1 acts through directly activating the conserved mitosis blocker WEE1, which is sufficient to bypass the LMI1 requirement for leaf proportionality.
Keywords: leaf development; live imaging; organ proportions; plant homeobox.
© 2018 Vuolo et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Bar M, Ori N. 2014. Leaf development and morphogenesis. Development 141: 4219–4230. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials