

Collagen prolyl 4-hydroxylase 1 is essential for HIF-1α stabilization and TNBC chemoresistance
- PMID: 30367042
- PMCID: PMC6203834
- DOI: 10.1038/s41467-018-06893-9
Collagen prolyl 4-hydroxylase 1 is essential for HIF-1α stabilization and TNBC chemoresistance
Abstract
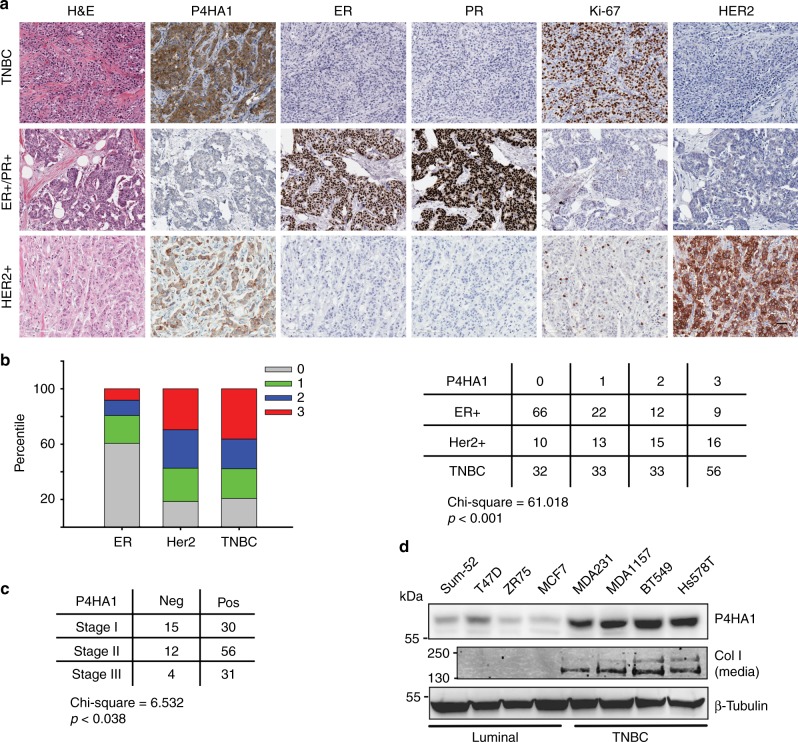
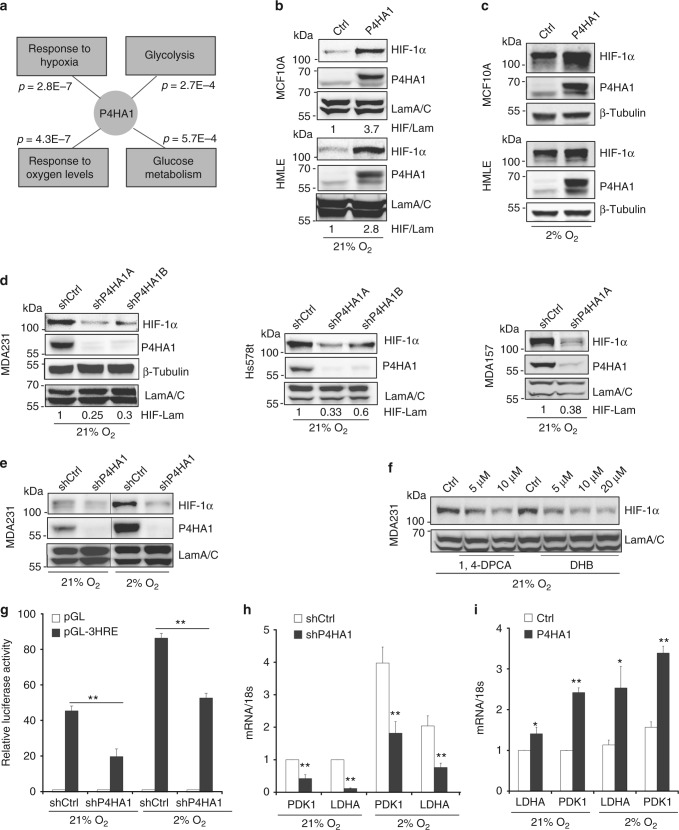
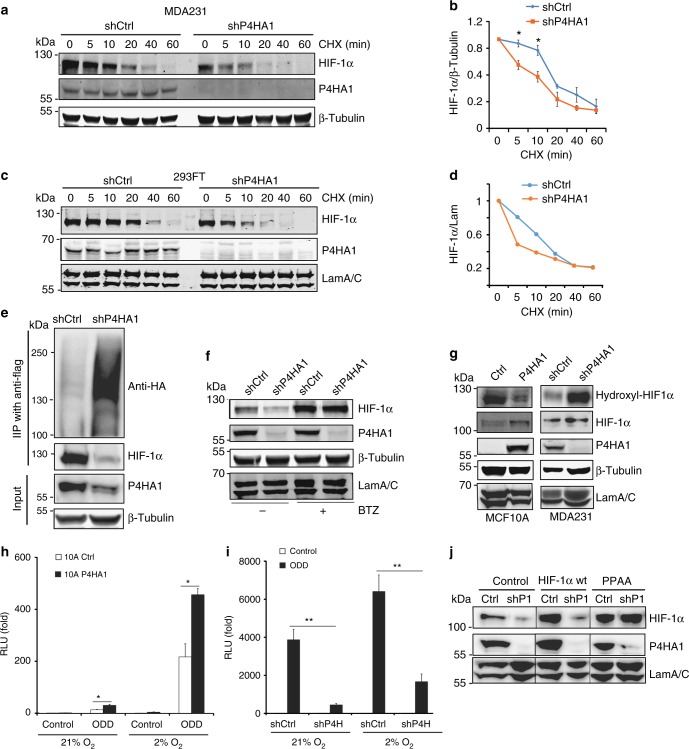
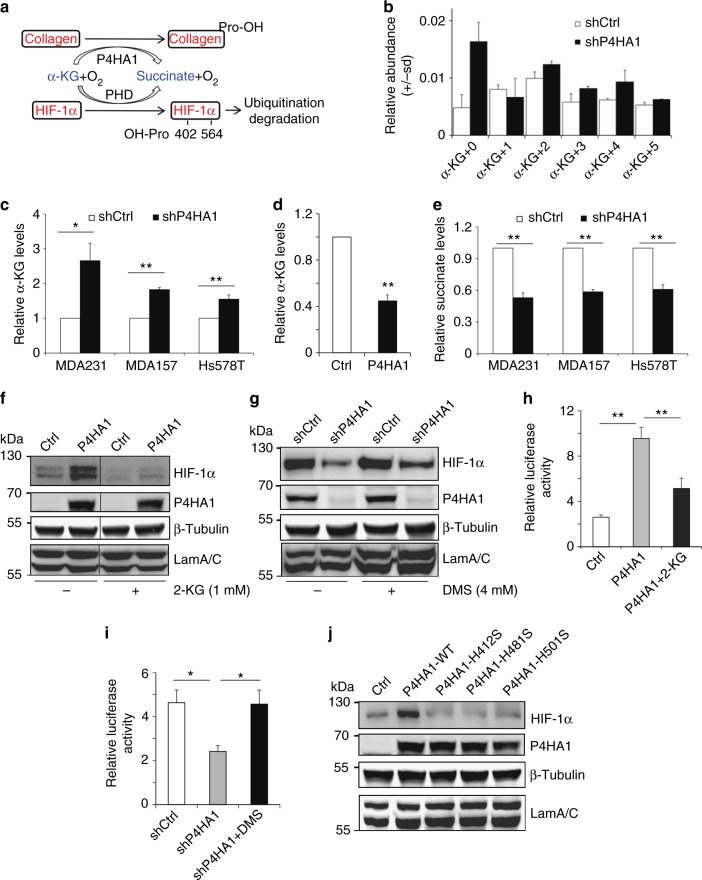
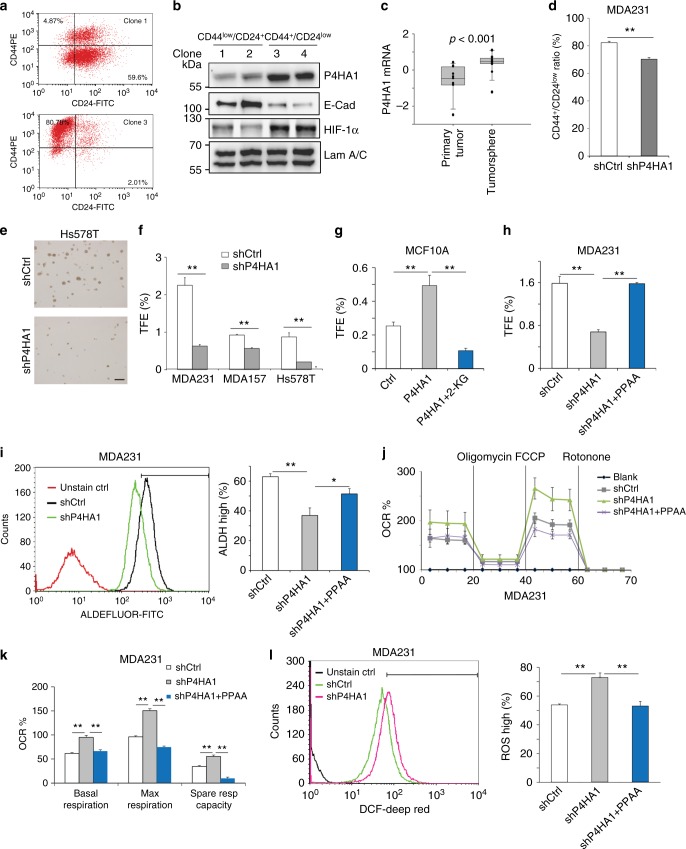
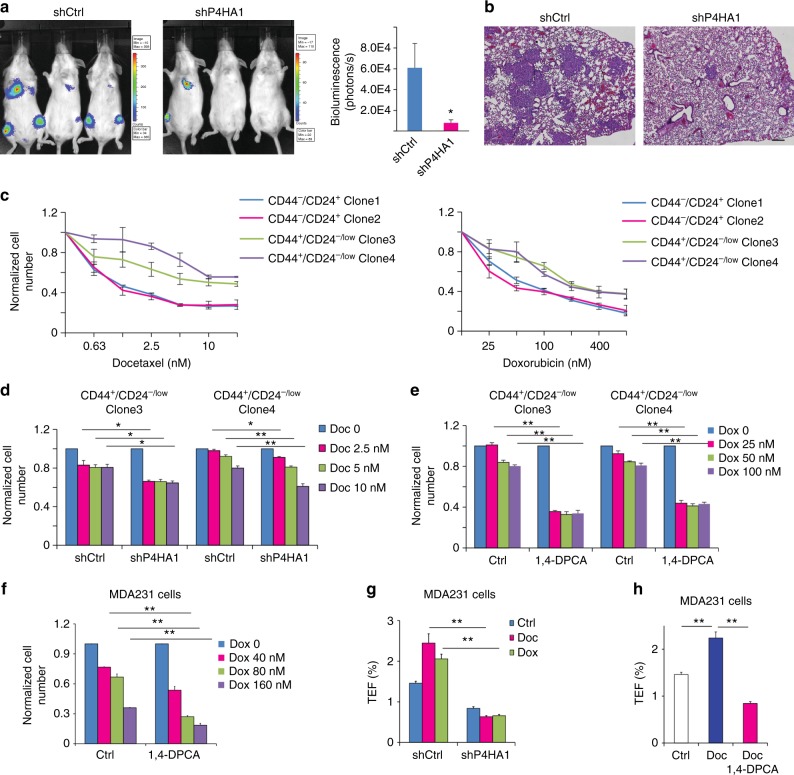
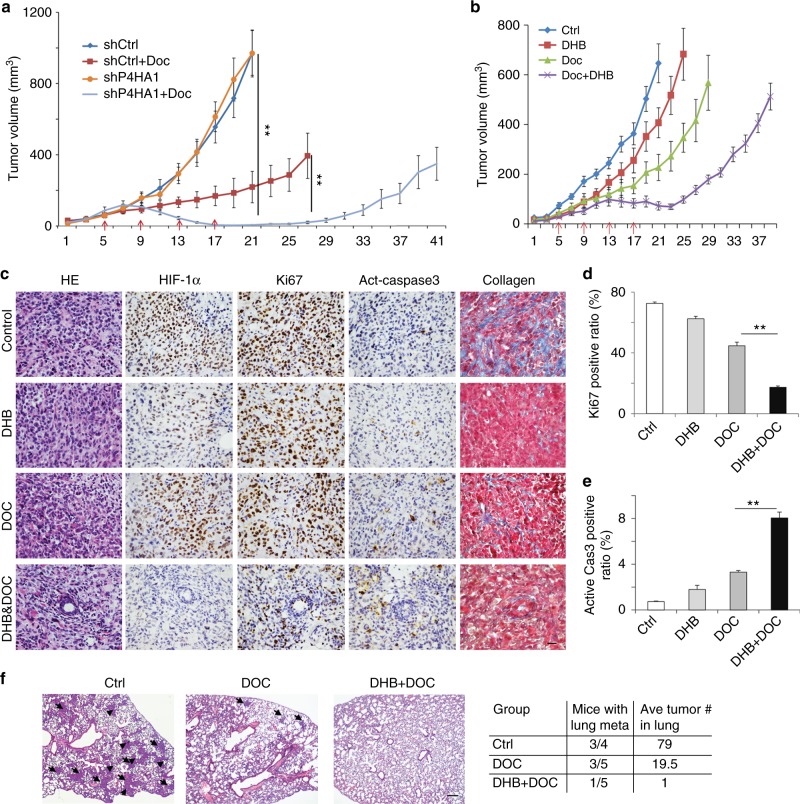
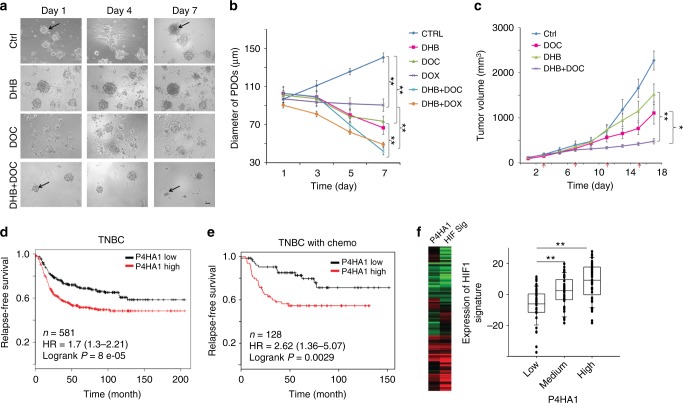
Collagen prolyl 4-hydroxylase (P4H) expression and collagen hydroxylation in cancer cells are necessary for breast cancer progression. Here, we show that P4H alpha 1 subunit (P4HA1) protein expression is induced in triple-negative breast cancer (TNBC) and HER2 positive breast cancer. By modulating alpha ketoglutarate (α-KG) and succinate levels P4HA1 expression reduces proline hydroxylation on hypoxia-inducible factor (HIF) 1α, enhancing its stability in cancer cells. Activation of the P4HA/HIF-1 axis enhances cancer cell stemness, accompanied by decreased oxidative phosphorylation and reactive oxygen species (ROS) levels. Inhibition of P4HA1 sensitizes TNBC to the chemotherapeutic agent docetaxel and doxorubicin in xenografts and patient-derived models. We also show that increased P4HA1 expression correlates with short relapse-free survival in TNBC patients who received chemotherapy. These results suggest that P4HA1 promotes chemoresistance by modulating HIF-1-dependent cancer cell stemness. Targeting collagen P4H is a promising strategy to inhibit tumor progression and sensitize TNBC to chemotherapeutic agents.
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
P4HA1 is a new regulator of the HIF-1 pathway in breast cancer.Cell Stress. 2019 Jan 9;3(1):27-28. doi: 10.15698/cst2019.01.173. Cell Stress. 2019. PMID: 31225497 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA223164/CA/NCI NIH HHS/United States
- R01 CA215095/CA/NCI NIH HHS/United States
- W81XWH-15-1-0052/DOD | Congressionally Directed Medical Research Programs (CDMRP)/International
- CA207772, CA215095, CA209045/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)/International
- R01 CA109136/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
