

Selection, hybridization, and the evolution of morphology in the Lake Malaŵi endemic cichlids of the genus Labeotropheus
- PMID: 30367138
- PMCID: PMC6203788
- DOI: 10.1038/s41598-018-34135-x
Selection, hybridization, and the evolution of morphology in the Lake Malaŵi endemic cichlids of the genus Labeotropheus
Abstract
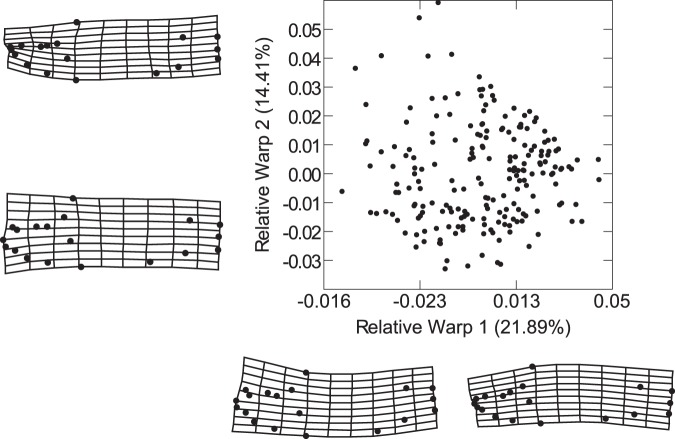
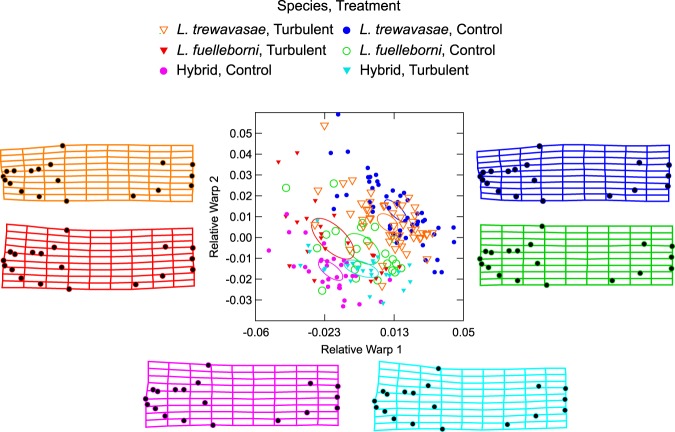
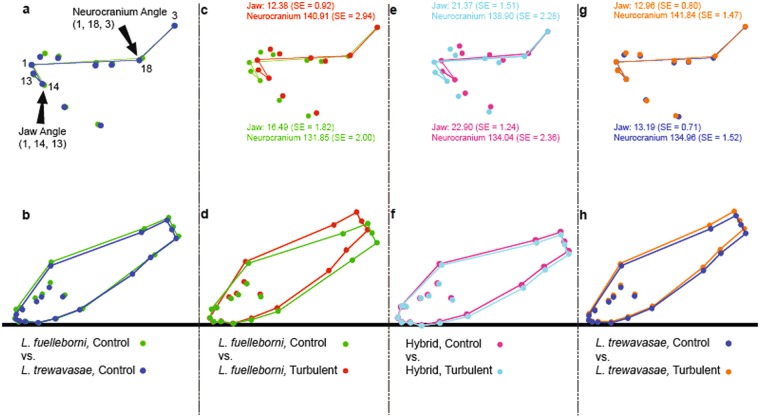
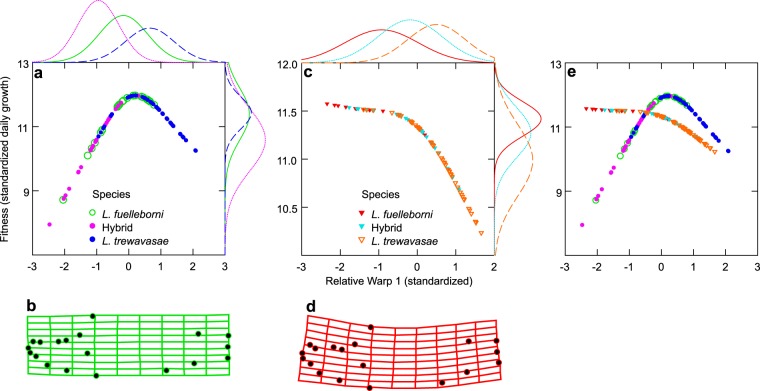
The cichlid fishes of Lake Malaŵi are the paramount example of adaptive radiation in vertebrates. Evidence of their astounding diversity is perhaps most visible in their adaptations for obtaining food; the genus Labeotropheus, due to their prominent snouts, are an interesting example of an extreme adaptation for feeding. Two different body types are found in this genus: a deep-bodied form (e.g., L. fuelleborni) found most often in turbulent shallow water; and a slender bodied form (e.g., L. trewavasae) found in structurally-complex deep water habitats. Here we test the hypothesis that L. trewavasae should suffer a loss in fitness, measured as growth rate, if raised in turbulence; additionally, we examined growth and morphology of L. fuelleborni and L. fuelleborni x L. trewavasae hybrids under these conditions. We did find the predicted loss of fitness in turbulent-raised L. trewavasae, but found no loss of fitness for L. fuelleborni in either condition; hybrids, due to an unusual morphology, performed better in turbulent as opposed to control conditions. Fitness in turbulent conditions was dependent upon morphology, with deeper bodies and upturned neurocrania allowing a greater growth rate under these conditions. Directional selection on morphology was crucial in the evolution of morphology in the Labeotropheus.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- FRYER GEOFFREY. THE TROPHIC INTERRELATIONSHIPS AND ECOLOGY OF SOME LITTORAL COMMUNITIES OF LAKE NYASA WITH ESPECIAL REFERENCE TO THE FISHES, AND A DISCUSSION OF THE EVOLUTION OF A GROUP OF ROCK-FREQUENTING CICHLIDAE. Proceedings of the Zoological Society of London. 2009;132(2):153–281. doi: 10.1111/j.1469-7998.1959.tb05521.x. - DOI
-
- Fryer, G., & Iles, T. D. The Cichlid Fishes of the Great Lakes of Africa (Oliver and Boyd, 1972).
-
- Ribbink AJ, Marsh AC, Marsh BA, Sharp BJ. The zoogeography, ecology and taxonomy of the genus Labeotropheus Ahl, 1927, of Lake Malawi (Pisces: Cichlidae) Zool. J. Linn. Soc. 1983;79:223–43. doi: 10.1111/j.1096-3642.1983.tb01166.x. - DOI
-
- Reinthal PN. The feeding habits of a group of herbivorous rock-dwelling cichlid fishes (Cichlidae, Perciformes) from Lake Malawi, Africa. Env. Biol. Fish. 1990;27:215–233. doi: 10.1007/BF00001674. - DOI
-
- Ribbink, A. J. Distribution and ecology of the cichlids of the African Great Lakes in (ed. Keenleyside, M. H. A.) Cichlid Fishes: Behaviour, Ecology and Evolution 36–59 (Chapman & Hall, 1991).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
