

IL-7 receptor blockade blunts antigen-specific memory T cell responses and chronic inflammation in primates
- PMID: 30367166
- PMCID: PMC6203796
- DOI: 10.1038/s41467-018-06804-y
IL-7 receptor blockade blunts antigen-specific memory T cell responses and chronic inflammation in primates
Abstract
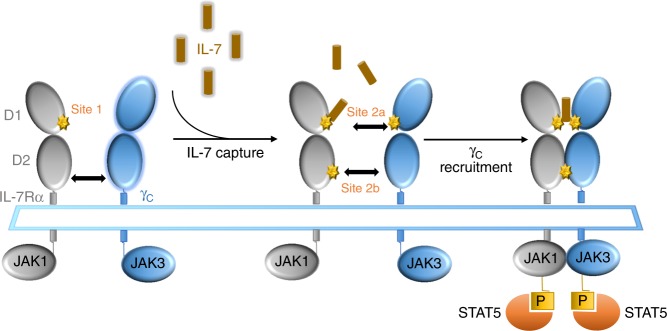
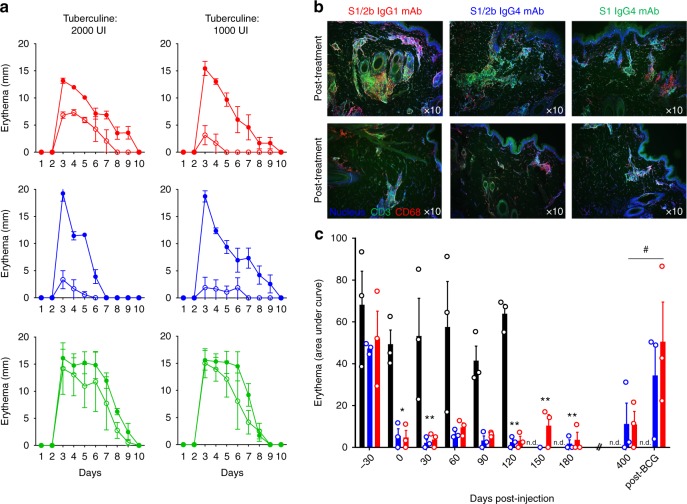
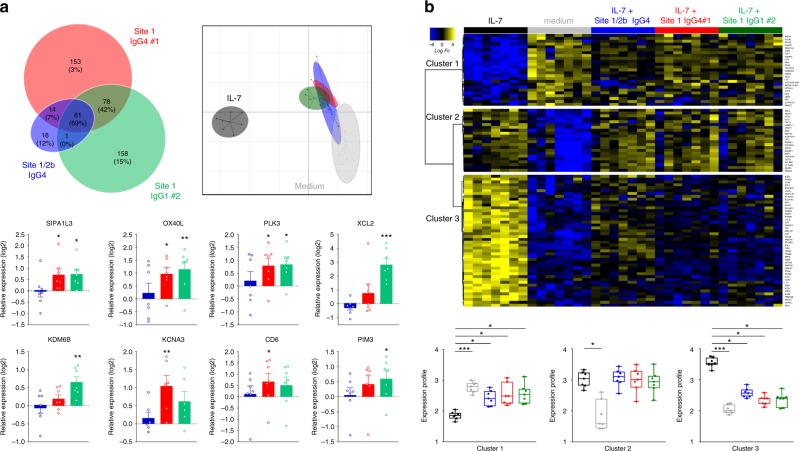
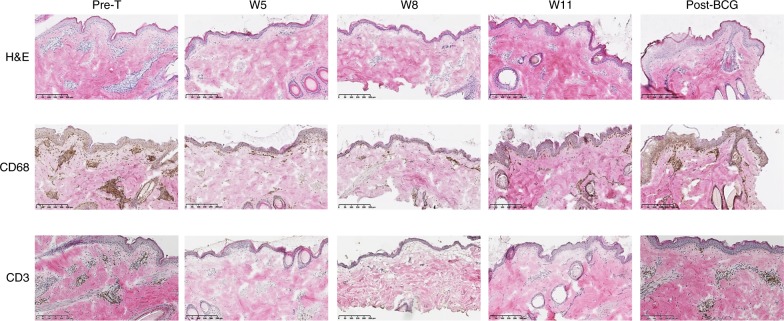
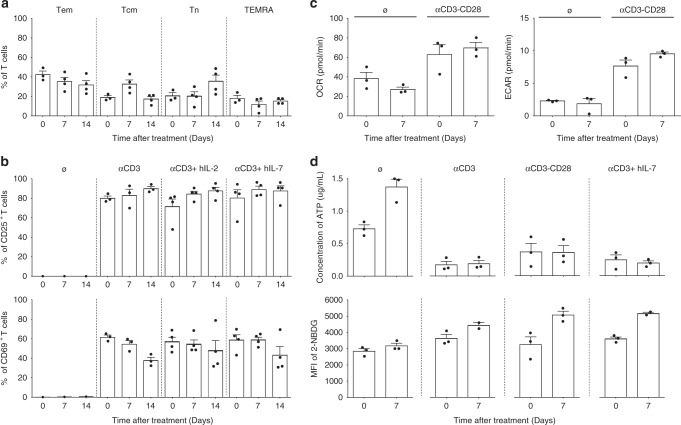
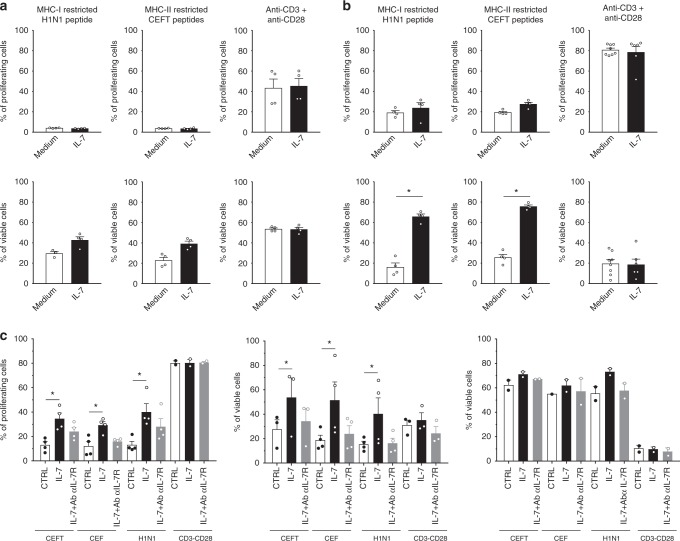
Targeting the expansion of pathogenic memory immune cells is a promising therapeutic strategy to prevent chronic autoimmune attacks. Here we investigate the therapeutic efficacy and mechanism of new anti-human IL-7Rα monoclonal antibodies (mAb) in non-human primates and show that, depending on the target epitope, a single injection of antagonistic anti-IL-7Rα mAbs induces a long-term control of skin inflammation despite repeated antigen challenges in presensitized monkeys. No modification in T cell numbers, phenotype, function or metabolism is observed in the peripheral blood or in response to polyclonal stimulation ex vivo. However, long-term in vivo hyporesponsiveness is associated with a significant decrease in the frequency of antigen-specific T cells producing IFN-γ upon antigen restimulation ex vivo. These findings indicate that chronic antigen-specific memory T cell responses can be controlled by anti-IL-7Rα mAbs, promoting and maintaining remission in T-cell mediated chronic inflammatory diseases.
Conflict of interest statement
The authors of this manuscript have conflicts of interest to disclose: CM, JPS, SB, BV and NP are shareholders of OSE Immunotherapeutics, a company owning patented anti-IL-7 receptor antagonists in clinical development. The remaining authors declare no competing interests.
Figures


Comment in
-
Full antagonist of the IL-7 receptor suppresses chronic inflammation in non-human primate models by controlling antigen-specific memory T cells.Cell Stress. 2018 Dec 10;2(12):362-364. doi: 10.15698/cst2018.12.168. Cell Stress. 2018. PMID: 31225460 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
